
Desde o famoso caso de Phineas Gage, lesões do lobo frontal têm sido associadas ao desenvolvimento de comportamento antissocial impulsivo. Este caso é ilustrativo a ponto de justificar uma breve descrição da sua apresentação clínica: Phineas Gage trabalhava na construção de estradas de ferro nos Estados Unidos, em meados do século XIX. Era descrito como equilibrado, meticuloso e persistente quanto aos seus objetivos, além de profissional responsável e habilidoso. Em um acidente nas explosões de rotina para abertura de túneis nas rochas da região, Phineas Gage foi atingido por uma barra de ferro que transpassou seu cérebro, entrando pela face esquerda, abaixo da órbita, e saindo pelo topo da cabeça. Surpreendentemente, Phineas Gage permaneceu consciente após o acidente, sobreviveu às esperadas infecções no seu ferimento e dois meses após o acidente estava recuperado, sem déficits motores e com linguagem e memória preservadas. A sua personalidade, no entanto, havia se modificado completamente. Phineas Gage transformou-se em uma pessoa impaciente, com baixo limiar à frustração, desrespeitoso com as outras pessoas, incapaz de adequar-se às normais sociais e de planejar o futuro. Não conseguiu estabelecer vínculos afetivos e sociais duradouros novamente ou fixar-se em empregos (Damásio, 1994).
A partir do infortúnio de Phineas Gage, relatos de caso e estudos retrospectivos de veteranos de guerra vêm mostrando a associação entre lesões pré-frontais – mais especificamente lesões nas porções ventromediais do córtex frontal – e a observação clínica de comportamento impulsivo, agressividade, jocosidade e inadequação social (Brower e Price, 2001). “Sociopatia adquirida” é o termo que tem sido frequentemente utilizado para descrever a mudança de personalidade observada em decorrência de danos cerebrais em regiões pré-frontais. Esses dados levaram à sugestão de que um comprometimento do funcionamento do lobo frontal ventromedial poderia contribuir para problemas relacionados ao controle de impulso e personalidade antissocial (Damásio, 2000). A variedade de déficits neuropsicológicos descritos em antissociais (Morgan e Lilienfeld, 2000) estaria em consonância com esta hipótese.
Estudos de neuroimagem
Os estudos de neuroimagem estrutural com ressonância nuclear magnética apontam alterações volumétricas do lobo frontal no TPAS. Comparando pacientes com diagnóstico de TPAS com controles não clínicos, pacientes com dependência de substâncias psicoativas e pacientes com outros diagnósticos psiquiátricos, Raine et al. (2000) verificaram que os pacientes com TPAS apresentavam uma redução do volume da matéria cinzenta pré-frontal e que esta redução correlacionava-se com uma diminuição da resposta autonômica a um evento estressor provocado experimentalmente – no caso, a realização de um discurso.
A amígdala é outra estrutura que estudos volumétricos tem implicado na fisiopatogenia do TPAS. Tiihonen et al.(2001) verificaram que o volume da amígdala correlacionou-se negativamente com os escores do PCL-R em criminosos violentos. Também foi descrita uma associação entre escores elevados no PCL-R e reduções bilaterais do volume de hipocampo posterior em criminosos violentos (Laasko et al., 2001). Esses últimos resultados devem ser tomados com cuidado, por se tratarem de amostra pequena, com comorbidade com dependência ao álcool e sem grupo-controle.
Há ainda indícios do envolvimento de outras estruturas cerebrais na ocorrência de TPAS. Em um estudo publicado mais recentemente, verificou-se que pacientes antissociais, comparados com controles saudáveis, apresentavam várias anormalidades no corpo caloso, o que poderia ser considerado como sugestivo de alterações no neurodesenvolvimento (Raine et al., 2003).
Os avanços em técnicas de neuroimagem funcional, como tomografia por emissão de pósitrons (PET), tomografia computadorizada por emissão de fóton único (SPECT) e ressonância magnética funcional (fMRI), permitiram que as relações entre região cerebral e diagnóstico específico e/ou processos mentais específicos fossem exploradas de maneira mais minuciosa.
Os estudos com PET (Goyer et al., 1994; Wong et al., 1997; Raine et al., 1994; 1998) e SPECT (Amen et al., 1996) também indicam o envolvimento de córtex pré-frontal no comportamento antissocial, com vários estudos demonstrando redução do metabolismo em regiões frontais. Em artigo de revisão considerando os artigos publicados de 1966 a 2000, Bassarath (2001) concluiu que estudos funcionais realizados até aquele momento (PET e SPECT) permitiam classificar como “robusto” o envolvimento do córtex pré-frontal, especialmente regiões mediais e laterais, no comportamento antissocial. Além do lobo frontal, também têm sido descritas reduções do metabolismo em estruturas subcorticais do sistema límbico (Amen et al., 1996), amígdala (Raine et al., 1997), hipocampo e núcleo caudado (Soderstrom et al., 2002).
Estudos mais recentes, utilizando-se de técnicas de fMRI, também apontam na direção do envolvimento de regiões pré-frontais e do sistema límbico no TPAS. Kiehl et al. (2001) demonstraram que criminosos psicopatas, comparados com criminosos não-psicopatas e controles sãos, apresentavam uma atenuação da ativação do complexo amígdala-hipocampo, giro paraipocampal, estriado ventral e giro do cíngulo posterior e anterior durante o processamento de palavras de valência negativa.
O comprometimento dos mecanismos envolvidos na aquisição de medo condicionado também tem sido implicado na fisiopatogenia do TPAS. Utilizando-se de uma tarefa baseada em teorias do condicionamento clássico (faces neutras pareadas com odor aversivo), Schneider et al. (2000) verificaram que pacientes com TPAS não diferiam de controles saudáveis quanto à aquisição de condicionamento, inferido a partir da observação do comportamento e medidas subjetivas. No entanto, observou-se que os pacientes apresentavam um aumento da intensidade de sinal na amígdala e no córtex pré-frontal dorsolateral, indo em direção oposta aos controles. Os autores explicaram este resultado argumentando que os pacientes necessitariam de um esforço adicional para o processamento de emoções negativas.
A possível necessidade de esforço adicional também foi observada em pacientes com TPAS e TPB durante a realização de um paradigma de inibição de comportamento estabelecido, denominado Go/No-Go. Ativações de córtex dorsolateral e orbitofrontal, especialmente à direita, durante a inibição comportamental têm sido consistentemente replicadas em voluntários saudáveis com esse paradigma. Pacientes com TPAS não diferiram de controles quanto ao desempenho na tarefa, mas apresentaram ativações mais extensas, envolvendo inclusive hemisfério esquerdo de córtex frontal medial e inferior, cíngulo anterior e regiões temporais (Vollm et al., 2004). Estes resultados foram explicados como uma estratégia compensatória, ou seja, o sucesso no desempenho da tarefa dependeria do recrutamento de áreas cerebrais adicionais.
Alguns resultados, no entanto, não confirmam a hipótese da necessidade de esforços compensatórios. Veit et al.(2002), utilizando um paradigma de condicionamento aversivo bastante semelhante ao descrito anteriormente (faces pareadas com pressão dolorosa ao invés de odor aversivo), obtiveram resultados opostos ao previsto pela hipótese. Psicopatas apresentavam ativações menos pronunciadas e mais breves em córtex orbitofrontal, ínsula, cíngulo anterior e amígdala, em comparação com controles saudáveis durante a execução da tarefa.
Os dados de neuroimagem claramente apontam para o envolvimento de algumas estruturas cerebrais específicas no desenvolvimento de comportamento antissocial. No entanto, algumas limitações metodológicas devem ser levadas em consideração na comparação dos resultados obtidos até o momento, bem como na sua extrapolação e generalização. Diferenças conceituais e de nomenclatura podem interferir nos resultados, havendo a necessidade de estudos com grupos mais homogêneos, sem comorbidades e com foco claro em comportamentos mais específicos (Séguin 2004). Além disso, deve-se lembrar também que boa parte dos estudos foi realizada com criminosos violentos, e que as alterações até então descritas poderiam relacionar-se de maneira mais específica com impulsividade, agressividade ou mesmo encarceramento, e não com a condição mais ampla de sociopatia. A aplicação de paradigmas específicos de ativação psicológica em voluntários saudáveis tem sido utilizada como um passo intermediário para a exploração de maneira sistemática dos diferentes componentes psíquicos possivelmente envolvidos no comportamento antissocial (exemplos em Moll et al., 2002; Anderson et al., 2002).
Estudos dos sistemas de neurotransmissão
Pode-se dizer, de maneira bastante simplificada, que a investigação sobre o papel de um determinado neurotransmissor em funções fisiológicas e patológicas poderia ser feita por meio do uso de drogas que ativam ou bloqueiam receptores específicos, pré e/ou pós-sinápticos. Além disso, a averiguação de seus precursores e metabólitos também seria ferramenta útil para a exploração da função de determinado neurotransmissor. A figura 1 ilustra os diferentes processos envolvidos na sinapse serotonérgica.
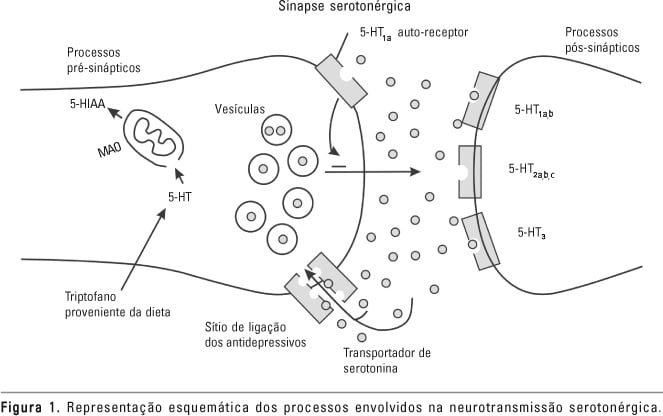
O prejuízo da função serotonérgica (5-HT) tem sido implicado na etiologia de vários transtornos mentais, entre eles transtornos de ansiedade, depressão e transtornos relacionados ao controle do impulso. Isso se deve a alguns achados comuns a todos, ou pelo menos a parte desses transtornos, como redução na concentração no líquido cefalorraquidiano (LCR) do produto final do metabolismo de 5-HT, o ácido 5-hidroxi-indolacético (5-HIAA), respostas hormonais atenuadas a desafios farmacológicos com drogas que interferem na função serotonérgica e alterações nas concentrações de receptores serotonérgicos (Mann et al., 1989, 1995; O’Keane et al., 1992; Coccaro et al., 1995; Dolan et al., 2001; Drevets et al., 1999). Além disso, os inibidores seletivos da recaptação de serotonina têm se mostrado efetivos no tratamento desses transtornos, o que se acredita ser devido ao aumento da neurotransmissão 5-HT (Bell e Nutt,1998; Mann et al., 2001; Jetty et al., 2001; Nemeroff, 2002).
Especificamente na TPAS, vários estudos também têm sugerido a ocorrência de anormalidades no funcionamento serotonérgico, especialmente no caso de criminosos violentos. A associação entre redução da função serotonérgica (5-HT) e comportamento agressivo e impulsivo tem sido demonstrada tanto em animais (Cherek e Lane, 1999) como em populações com diagnóstico de personalidade antissocial (Fairbanks et al., 2001; Dolan et al., 2001).
Em voluntários saudáveis, a depleção aguda de triptofano, um aminoácido proveniente da dieta e precursor de 5-HT, induziu aumento da velocidade de processos psicomotores, mas tornou a escolha do comportamento mais lenta (Rogers et al., 1999a). Os autores concluíram que a serotonina normalmente reduz a velocidade de processos psicomotores nos circuitos dorso-fronto/estriatal e que o prejuízo da função serotonérgica seria um dos mecanismos de resposta impulsiva.
Criminosos antissociais e violentos apresentaram níveis plasmáticos significativamente mais elevados de triptofano livre que controles saudáveis, sugerindo um distúrbio do metabolismo de triptofano na fisiopatogenia da sociopatia (Tiihonen et al., 2001). Este mesmo grupo de pesquisadores sugeriu, a partir do estudo do caso de um jovem de 15 anos com diagnóstico de transtorno de conduta, que os níveis elevados de triptofano poderiam ser um indicador precoce de comportamento criminoso no futuro (Virkkunen et al., 2003).
Outra medida da associação entre prejuízo do funcionamento das vias serotonérgicas e comportamento antissocial é a diminuição das concentrações do 5-HIAA no LCR de criminosos impulsivos, demonstrada em diferentes estudos (Brown et al., 1982, Soderstrom et al., 2003; Constantino et al., 1997; Coccaro et al., 1990). Os baixos níveis de 5-HIAA no LCR sugeririam que o déficit estaria na liberação de serotonina, mas, por outro lado, a estimulação direta de receptores pós-sinápticos do tipo 5-HT2, por meio de desafios farmacológicos, também mostraram respostas alteradas.
Os desafios farmacológicos se caracterizam pela administração aguda de drogas provocadoras de aumento da função serotonérgica cerebral, que, por sua vez, pode levar a um aumento na secreção de alguns hormônios pituitários, como prolactina e corticotrofina (ACTH). As mudanças observadas nos níveis hormonais plasmáticos em decorrência da administração aguda de drogas têm sido frequentemente usadas, tanto em voluntários saudáveis quanto em pacientes psiquiátricos, como uma medida indireta da função serotonérgica no sistema nervoso central (Cowen, 1998).
A administração aguda do agonista serotonérgico m-cloro-fenilpiperazina (mCPP) em indivíduos com diagnóstico de TPAS resultou em respostas hormonais alteradas, quando comparados com voluntários saudáveis (Moss et al., 1990). O aumento dos níveis de cortisol foi significativamente mais pronunciado nos pacientes que nos controles, enquanto os níveis de prolactina apresentaram resposta em direção oposta, sendo mais atenuados nos pacientes. A administração aguda de d-fenfluramina, um inibidor da recaptação e potente liberador de 5-HT, levou a aumentos menos pronunciados dos níveis plasmáticos de prolactina em criminosos com diagnóstico de TPAS, comparado com a resposta de controles sãos (O´Keane et al., 1992).
Recentemente, aplicando o conceito mais amplo de psicopatia e seus diferentes componentes combinados com um desafio farmacológico com d-fenfluramina em criminosos violentos, Dolan e Anderson (2003) verificaram que traços impulsivos de personalidade correlacionavam-se negativamente com a função serotonérgica, enquanto traços de arrogância correlacionavam-se positivamente, sendo este último dado interpretado como um possível componente adaptativo da psicopatia.
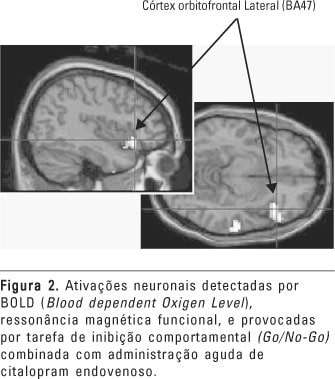
Embora os resultados de estudos com avaliação de respostas neuroendócrinas a testes provocativos farmacológicos sejam consistentes, eles não permitem que sejam localizadas as regiões do cérebro onde se encontrariam as deficiências neuroquímicas. O uso de neuroimagem para monitorar as respostas metabólicas ou fluxo sanguíneo para testes provocativos farmacológicos poderia ser um meio mais adequado para avaliar a responsividade farmacológica por regiões cerebrais específicas. Estudos com PET e d-fenfluramina mostraram aumento da atividade metabólica no córtex frontal ventromedial em voluntários normais e uma falta de resposta em pacientes agressivos e impulsivos com TBP (Siever et al., 1999; Soloff et al., 2000). A administração endovenosa de mCPP em voluntários normais e em pacientes com dependência ao álcool causou ativação em várias áreas do córtex pré-frontal (Hommer et al., 1997). Em voluntários saudáveis, o mCPP acentuou a resposta neuronal, detectada por fMRI, de regiões laterais do córtex orbitofrontal durante a execução de tarefa de inibição de comportamento (Go/No-Go) (Anderson et al., 2002). Efeitos semelhantes foram obtidos com a administração endovenosa de citalopram, um inibidor seletivo da recaptação de serotonina (Del-Ben et al., 2003). Os efeitos do citalopram nesta tarefa estão ilustrados na figura 2. Esses resultados sugerem um papel da serotonina na inibição de comportamentos, por meio da facilitação de mecanismos frontais.
Outros sistemas de neurotransmissão também têm sido associados a comportamento antissocial. Uma hipótese intrigante implica os mecanismos envolvidos no processamento de recompensa, e consequentemente o sistema dopaminérgico, na fisiopatogenia do TPAS (Blum et al., 2000). A hipótese se aplicaria também a outros transtornos mentais, como dependência química e transtornos do controle do impulso, relacionando-os a uma ruptura na cascata de respostas a estímulos de recompensa. O funcionamento do sistema dopaminérgico estaria diminuído devido a um comprometimento de receptores pós-sinápticos do tipo D2.
Hipóteses a respeito da fisiopatogenia do transtorno de personalidade antissocial
Resumidamente, os dados clínicos obtidos até o momento sugerem que, do ponto de vista anatômico, porções ventromediais do lobo frontal, particularmente o córtex orbitofrontal (COF), e outras estruturas do sistema límbico, especialmente a amígdala, estariam envolvidas na patogênese do TPAS. Além disso, redução da função serotonérgica também estaria relacionada com TPAS, se não com todos os seus aspectos, pelo menos com o seu componente impulsivo/agressivo.
Estudos experimentais indicam que o processamento de estímulos de recompensa e punição, bem como a escolha entre as opções de comportamento possíveis frente às características do reforço apresentado (magnitude, probabilidade de ocorrência, tempo de latência), são essencialmente mediados por circuitaria neural que inclui as estruturas anatômicas supostamente implicadas na patogenia do TPAS, como o COF e a amígdala (Rolls 1999; Tremblay e Schultz 1999; Schoenbaum et al., 1998). Pode-se supor que, no TPAS, as representações de respostas não evocariam as representações de reforço normalmente esperadas; o comportamento passaria, então, a ser governado por mecanismos mais primitivos e menos precisos.
A inibição de comportamentos punidos também seria mediada pelo COF. Regiões laterais deste parecem estar particularmente envolvidas em tarefas que requerem inibição de respostas motoras preponderantes (Elliott et al., 2000). Do ponto de vista adaptativo, mecanismos inibitórios podem ser cruciais para o funcionando social normal. Blair et al. (1999) demonstraram ativação de COF lateral para expressões faciais de raiva, que representariam um sinal social para inibir comportamento inapropriado. Assim, a deterioração em funções do COF lateral poderia contribuir para comportamentos impulsivos ou desinibidos, frequentemente observados em pacientes com TPAS.
Por outro lado, as regiões mediais do COF mediariam a escolha do comportamento por meio da representação das consequências motivacionais das escolhas. De acordo com Damasio et al. (1990), as porções mediais do COF processariam a escolha da resposta comportamental frente a estímulos arriscados, especialmente em situações sociais, desencadeando como sinal de alerta sensações somáticas ou viscerais (gut feeling). Essa região seria essencial para o planejamento do futuro e tomada de decisões. Pacientes com lesões em regiões ventromediais do COF apresentam respostas autonômicas atenuadas e fazem escolhas arriscadas em um paradigma experimental que simula um jogo de azar (gambling) (Bechara et al., 1996). Reciprocamente, sujeitos saudáveis mostram ativação dessa mesma região do COF por um paradigma semelhante (Rogers et al., 1999b).
Os estudos animais sugerem que a inervação serotonérgica do COF, da amígdala e do estriado ventral seria ativada por meio de ameaças externas ou mecanismos de ansiedade antecipatória. A via serotonérgica ascendente proveniente do núcleo dorsal da rafe mediaria respostas adaptativas a situações adversas atuais ou futuras por meio de receptores pós-sinápticos do tipo 5-HT2, a serotonina inibiria comportamentos de aproximação, mediados por dopamina, e facilitaria a evitação (Deakin e Graeff, 1991; Deakin, 2003). O COF tem projeções extensas para os centros de controle autonômico no hipotálamo medial e na matéria cinzenta periaquedutal, que parece mediar respostas do tipo luta-fuga (Graeff et al., 1997). A ansiedade induzida por esta resposta pode ser um caso especial de ansiedade antecipatória facilitada por 5-HT (Deakin e Graeff 1991; Deakin 1999). Assim, defeitos na função 5-HT e danos do COF podem resultar em falta de inibição, tanto de comportamento socialmente arriscado como punido.
Considerações finais
O interesse crescente no estabelecimento das bases neurais do comportamento antissocial que se observa atualmente provavelmente se deve, pelo menos em parte, ao aumento significativo da criminalidade e violência urbana em diferentes partes do mundo. Os avanços metodológicos obtidos nas últimas décadas, como, por exemplo, as técnicas de investigação em neuroimagem, têm permitido que diferentes hipóteses sobre as bases neurobiológicas de diferentes transtornos mentais sejam sucessivamente testadas. A identificação de fatores de risco, tanto psicossociais como biológicos, para a ocorrência de comportamento antissocial seria de extrema utilidade para o desenvolvimento de abordagens efetivas de prevenção e intervenção. No entanto, apesar de muitos avanços terem sido alcançados nessa área, deve-se ter cautela na interpretação dos resultados obtidos até o momento, particularmente na sua extrapolação para outras esferas não médicas, como moral, ética ou jurídica. Uma eventual aplicação das informações a respeito das bases biológicas do transtorno de personalidade antissocial em outros campos do conhecimento exigiria, antes de qualquer coisa, uma reflexão ampla e profunda de diferentes áreas da sociedade.
Referências:
- AMEN, D.G. et al. – Brain SPECT findings and aggressiveness. Ann Clin Psychiatry 8(3):129-37, 1996.
- AMERICAN PSYCHIATRIC ASSOCIATION. – Diagnostical and statistical manual of mental disorder. 4th ed. Washington DC: American Psychiatric Association, 1994.
- ANDERSON, I.M. et al. – 5-HT(2C) receptor activation by m-chlorophenylpiperazine detected in humans with fMRI. Neuroreport 13(12):1547-51, 2002.
- BASSARATH, L. – Neuroimaging studies of antisocial behaviour. Can J Psychiatry 46(8):728-32, 2001.
- BECHARA, A. et al – Failure to respond autonomically to anticipated future outcomes following damage to prefrontal cortex. Cereb Cortex 6(2):215-25, 1996.
- BELL, C.J.; Nutt, D.J. – Serotonin and panic. Br J Psychiatry 172:465-71, 1998.
- BLAIR, R.J. et al – Dissociable neural responses to facial expressions of sadness and anger. Brain 122:883-93, 1999.
- BLAIR, R. J. – Neurobiological basis of psychopathy. Br J Psychiatry 182:5-7, 2003.
- BLAIR, R.J. – Neurocognitive models of aggression, the antisocial personality disorders, and psychopathy. J Neurol Neurosurg Psychiatry 71(6):727-31, 2001.
- BLUM, K.; BRAVERMAN, E.R.; HOLDER, J.M. et al. – Reward deficiency syndrome: a Biogenetic Model for the Diagnosis and Treatment of Impulsive, Addictive, and COmpulsive Behaviors. J Psychoactive Drugs. 32 )(suppl IV):1-112, 2000.
- BROWER, M.C.; PRICE, B.H. – Neuropsychiatry of frontal lobe dysfunction in violent and criminal behaviour:a critical review. J Neurol Neurosurg Psychiatry 71(6):720-6, 2001.
- BROWN, G.L. et al. – Aggression in humans correlates with cerebrospinal fuid amine metabolites Psychiatry Res1:131-9, 1979.
- CADORET, R.J.; STEWART, M.A. – An adoption study of attention deficit/hyperactivity/ aggression and their relationship to adult antisocial personality. Compr Psychiatr 32:73-82,1991.
- CLARKIN, J.F. et al. – The Personality Disorders Institute/Borderline Personality Disorder Research Foundation randomized control trial for borderline personality disorder: rationale, methods, and patient characteristics. J Personal Disord 18(1):52-72, 2004.
- COCCARO, E.F. et al. – Preliminary evidence of a serotonin (5-HT-1-like) component to the prolactin response to buspirone challenge in humans. Arch Gen Psychiatry 47(6):594-5, 1990.
- COCCARO, E.F.; KAVOUSSI, R.J.; HAUGER, R.L. – Physiological responses to d-fenfluramine and ipsapirone challenge correlate with indices of aggression in males with personality disorder. Int Clin Psychopharmacol 10:177-9, 1995.
- COID, J.W.; CORDESS, C. – Compulsory admission of dangerous psychopaths. BMJ 20: 1581-2, 1992.
- CONSTANTINO, J.N.; MORRIS, J.A.; MURPHY, D.L. – CSF 5-HIAA and family history of antisocial personality disorder in newborns. Am J Psychiatry 154(12):1771-3, 1997.
- COWEN, P.J. – Neuroendocrine challenge tests: what can we learn from them? In: Van de Kar, L.D. (ed) Methods in neuroendocrinology. Boca Raton: CRC Press, pp. 205-23, 1998.
- CHEN, T.J.; COMINGS, D.E. – Reward deficiency syndrome: a biogenetic model for the diagnosis and treatment of impulsive, addictive, and compulsive behaviors. J Psychoactive Drugs 32(Suppl:i-iv):1-112, 2000.
- CHEREK, D.R.; LANE, S.D. – Effects of d,l-fenfluramine on aggressive and impulsive responding in adult males with a history of conduct disorder. Psychopharmacology (Berl) 146(4):473-81, 1999.
- DAMASIO, A.; TRANEL, D.; DAMASIO, H. – Individuals with psychopathic behaviour caused by frontal damage fail to respond autonomically to social stimuli. Brain and Behaviour Research 41:81-94, 1990.
- DAMASIO, A.R. – A neural basis for sociopathy. Arch Gen Psychiatry 57:128-30, 2000.
- DAMASIO, A. R. Descartes’ error: Emotion, reason, and the human brain. New York:Grosset/Putnam, 1994.
- DEAKIN, J.F. – Depression and antisocial personality disorder: two contrasting disorders of 5HT function. J Neural Transm Suppl (64):79-93, 2003.
- DEAKIN, J.F.W. – Making sense of serotonin (5HT) and its role in common psychopathology. In: Tansella, M.; Thornicroft, G. (eds.) Common Mental Disorders in Primary Care. London: Routledge, p.p. 17-33, 1999.
- DEAKIN, J.F.W.; GRAEFF, F.G. – 5-HT and mechanisms of defense. J Psychopharmacol 5:305-15, 1991.
- DEL-BEN, C.M. et al. – Effects of citalopram on brain activation during cognitive tasks involving behavioural inhibition and reinforcement. J Psychopharmacology 17(3):A14, 2003.
- DOLAN, M.; ANDERSON, I.M.; DEAKIN, J.F. – Relationship between 5-HT function and impulsivity and aggression in male offenders with personality disorders.Br J Psychiatry 178:352-9, 2001.
- DOLAN, M.C.; ANDERSON, I.M. – The relationship between serotonergic function and the Psychopathy Checklist: Screening Version. J Psychopharmacol 7(2):216-22, 2003.
- DREVETS, W.C. et al. – PET imaging of serotonin 1A receptor binding in depression. Biol Psychiatry 46:1375-87, 1999.
- ELLIOTT, R.; FRISTON, K.J.; DOLAN, R.J. – Dissociable neural responses in human reward systems. J Neurosci20(16):6159-665, 2000.
- EVENDEN, J.L. – Varieties of impulsivity. Psychopharmacol (Berl) 146:348-61, 1999.
- FAIRBANKS, L.A. et al – Social impulsivity inversely associated with CSF 5-HIAA and fluoxetine exposure in vervet monkeys. Neuropsychopharmacology 24(4):370-8, 2001.
- GOYER, P.F. et al – Positron-emission tomography and personality disorders. Neuropsychopharmacology 0(1):21-8, 1994.
- GRAEFF, F.G.; VIANA, M.B.; MORA, P.O. – Dual role of 5-HT in defense and anxiety. Neurosci Biobehav Rev 21:791-9, 1997.
- HARE, R.D.; HART, S.D.; HARPUR, T.J. – Psychopathy and the DSM-IV criteria for antisocial personality disorder. J Abnorm Psychol 100(3):391-8, 1991.
- HOLMES, S.E.; SLAUGHTER, J.R.; KASHANI, J. – Risk factors in childhood that lead to the development of conduct disorder and antisocial personality disorder. Child Psychiatry Hum Dev 31(3):183-93, 2001.
- HOMMER, D. et al. – Effects of m-chlorophenylpiperazine on regional brain glucose utilization: a positron emission tomographic comparison of alcoholic and control subjects. J Neurosci 7(8):2796-806, 1997.
- JETTY, P.V.; CHARNEY, D.S.; GODDARD, A.W. – Neurobiology of generalized anxiety disorder. Psychiatr Clin North Am 24:75-97, 2001.
- KIEHL, K.A. et al. – Limbic abnormalities in affective processing by criminal psychopaths as revealed by functional magnetic resonance imaging. Biol Psychiatry 50(9):677-84, 2001.
- LAAKSO, M.P. et al. – Psychopathy and the posterior hippocampus. Behav Brain Res 118(2):187-93, 2001.
- MANN, J.J. et al. – Evidence for the 5-HT hypothesis of suicide. A review of post-mortem studies. Br J PsychiatrySuppl (8):7-14, 1989.
- MANN, J.J. et al. Blunted serotonergic responsivity in depressed inpatients. Neuropsychopharmacology 13:53-64, 1995.
- MANN, J.J.; BRENT, D.A.; ARANGO, V. – The neurobiology and genetics of suicide and attempted suicide: a focus on the serotonergic system. Neuropsychopharmacology 24:467-77, 2001.
- MASON, D.A., FRICK, P.J. – The heritability of antisocial behavior: a meta-analysis of twin and adoption studies. Journal of Psychopathology and Behavioral Assessment 16 (4):301-23, 1994.
- McGUFFIN, P.; THAPAR, A. – The genetics of personality disorder. Br J Psychiatry 160:12-23, 1992.
- MOLL, J. et al. – Functional networks in emotional moral and nonmoral social judgments. Neuroimage 16(3 Pt 1):696-703, 2002.
- MORGAN, A.B.; LILIENFELD, S.O. – A meta-analytic review of the relation between antisocial behavior and neuropsychological measures of executive function. Clin Psychol Rev 20(1):113-36, 2000.
- MOSS, H.B.; YAO, J.K.; PANZAK, G.L. – Serotonergic responsivity and behavioral dimensions in antisocial personality disorder with substance abuse. Biol Psychiatry 28(4):325-38, 1990.
- NEMEROFF, C.B. – Recent advances in the neurobiology of depression. Psychopharmacol Bull 36 Suppl 2:6-23, 2002.
- O’CONNELL, S. – Mindreading. An Investigation into How We Learn to Love and Lie. London: Arrow Books Ltd., 1998.
- O’KEANE, V. et al. – Blunted prolactin responses to d-fenfluramine in sociopathy. Evidence for subsensitivity of central serotonergic function. Br J Psychiatry 160:643-6, 1992.
- RAINE, A. et al. – Selective reductions in prefrontal glucose metabolism in murderers. Biol Psychiatry 36(6):365-73, 1994.
- RAINE, A. et al. – Prefrontal glucose deficits in murderers lacking psychosocial deprivation. Neuropsychiatry Neuropsychol Behav Neurol 11(1):1-7, 1998.
- RAINE, A. et al. – Reduced prefrontal gray matter volume and reduced autonomic activity in antisocial personality disorder. Arch Gen Psychiatry 57(2):119-27, 2000.
- RAINE, A. et al. – Corpus callosum abnormalities in psychopathic antisocial individuals. Arch Gen Psychiatry60(11):1134-42, 2003.
- RAINE, A.; BUCHSBAUM, M.; LACASSE, L. – Brain abnormalities in murderers indicated by positron emission tomography. Biol Psychiatry 42(6):495-508, 1997.
- ROBINS, L.N.; TIPP, J.; PRZYBECK, T. – Antisocial personality. In: Robins, L.N.; Reiger, D.A. (eds.) Psychiatric Disorders in America. New York: Free Press, p.p. 258-290, 1991.
- ROGERS, R.D. et al. – Tryptophan depletion impairs stimulus-reward learning while methylphenidate disrupts attentional control in healthy young adults: implications for the monoaminergic basis of impulsive behaviour. Psychopharmacol 146:482-91, 1999a.
- ROGERS, R.D. et al. – Choosing between small, likely rewards and large, unlikely rewards activates inferior and orbital prefrontal cortex. J Neurosci 20:9029-38, 1999b.
- ROLLS, E.T. et al. – Responses to the sensory properties of fat of neurons in the primate orbitofrontal cortex. J Neurosci 19(4):1532-40, 1999.
- SANISLOW, C.A., GRILO, C.M.; MCGLASHAN, T.H. – Factor analysis of the DSM-III-R borderline personality disorder criteria in psychiatric inpatients. Am J Psychiatry 157(10):1629-33, 2000.
- SCHNEIDER, F. et al. – Functional imaging of conditioned aversive emotional responses in antisocial personality disorder. Neuropsychobiology 42(4):192-201, 2000.
- SCHOENBAUM, G.; CHIBA, A.A.; GALLAGHER, M. – Orbitofrontal cortex and basolateral amygdala encode expected outcomes during learning. Nat Neurosci 1(2):155-9, 1998.
- SIEVER, L.J. et al. – D,l-fenfluramine response in impulsive personality disorder assessed with [18F]fluorodeoxyglucose positron emission tomography. Neuropsychopharmacology 20(5):413-23, 1999.
- SINGLETON, N. et al. – Psychiatric Morbidity Among Prisoners. London: Stationery Office, 1998.
- SODERSTROM, H. et al. – CSF studies in violent offenders. I. 5-HIAA as a negative and HVA as a positive predictor of psychopathy. J Neural Transm 108(7):869-78, 2001.
- SODERSTROM, H. et al. – Reduced frontotemporal perfusion in psychopathic personality. Psychiatry Res 114(2):81-94, 2002.
- SOLOFF, P.H. et al. – A fenfluramine-activated FDG-PET study of borderline personality disorder. Biol Psychiatry47(6):540-7, 2000.
- TIIHONEN, J. et al. – Free L-tryptophan plasma levels in antisocial violent offenders. Psychopharmacology (Berl)157(4):395-400, 2001.
- TREMBLAY, L.; SCHULTZ, W. – Relative reward preference in primate orbitofrontal cortex. Nature 398(6729):704-8, 1999.
- VEIT, R. et al. – Brain circuits involved in emotional learning in antisocial behavior and social phobia in humans. Neurosci Lett 328(3):233-6, 2002.
- VIRKKUNEN, M. et al. – Total plasma l-tryptophan, free l-tryptophan and competing amino acid levels in a homicidal male adolescent with conduct disorder. Acta Psychiatr Scand 08(3):244-6, 2003.
- VOLLM, B. et al. – Neurobiological substrates of antisocial and borderline personality disorder: preliminary results of a functional fMRI study. Crim Behav Ment Health 14(1):39-54, 2004.
- WONG, M. et al. – Repetitive and non-repetitive violent offending behaviour in male patients in a maximum security mental hospital-clinical and neuroimaging findings. Med Sci Law 37(2):150-60, 1997.
Fonte:
Neurobiologia do transtorno de personalidade antissocial
Revista de Psiquiatria Clínica
print version ISSN 0101-6083
Rev. psiquiatr. clín. vol.32 no.1 São Paulo 2005
Cristina Marta Del-Ben
http://dx.doi.org/10.1590/S0101-60832005000100004


