
Muitas vezes temos uma falsa impressão de que a evolução é uma marcha solitária de populações de uma espécie acumulando transformações ao longo do tempo. Porém, ao olharmos com maior detalhe percebemos que, na prática, ocorre o contrário. Existe uma forte conexão entre todas as formas de vida na Terra e, em alguns casos, essa conexão é significativamente estreita, fazendo com que as transformações de uma espécie criem pressões para transformações de outras espécies, levando a um processo de transformações mútuas entre espécies.
Esse mecanismo de interações recíprocas que levam a mudanças evolutivas concomitantes entre espécies é conhecido como coevolução. Esse processo ocorre geralmente entre espécies que apresentam comportamento ecológico próximo, exercendo pressões seletivas mútuas. Em outras palavras, coevolução é o processo onde duas ou mais espécies evoluem juntas, fruto de sua interação.
Nesse sentido, podemos definir dois processos de coevolução: a coevolução difusa e a coevolução restrita. O primeiro está relacionado com uns aspectos mais amplos, envolvendo múltiplas espécies, por exemplo, um ecossistema onde as espécies interagem entre si criando pressões seletivas recíprocas. Apesar da coevolução difusa ser extremamente plausível ela é muito difícil de ser analisada pela questão de envolver inúmeras variantes. Já a coevolução restrita está relacionada essencialmente entre duas (ou poucas) espécies, e por envolver um menor número de espécies apresenta maior facilidade em ser estudada. Nesse texto analisaremos alguns casos clássicos de coevolução restrita.
Um dos indícios de que duas espécies podem estar coevoluindo pode ser sua estreita interação ecológica, porém nem toda interação ecológica é fruto de coevolução. Da mesma forma que coadaptações, ou seja, espécies diferentes podem apresentar adaptações mútuas, o que poderia ser um indício de que ocorreu coevolução. Porém, não é uma evidencia definitiva, pois é possível surgir coadaptações a partir de processos independentes.
Uma das evidências que apontariam à ocorrência de coevolução é quando dois táxons apresentarem filogenias em imagem especular, ou seja, uma co-filogenia, nela é como se a árvore filogenética de um táxon fosse colocada na frente de um espelho. Na imagem abaixo vemos a filogenia de Tetraopes (besouro) norte-americano e sua planta alimentícia, as Asdepias, que é uma representação clássica de co-filogenia.
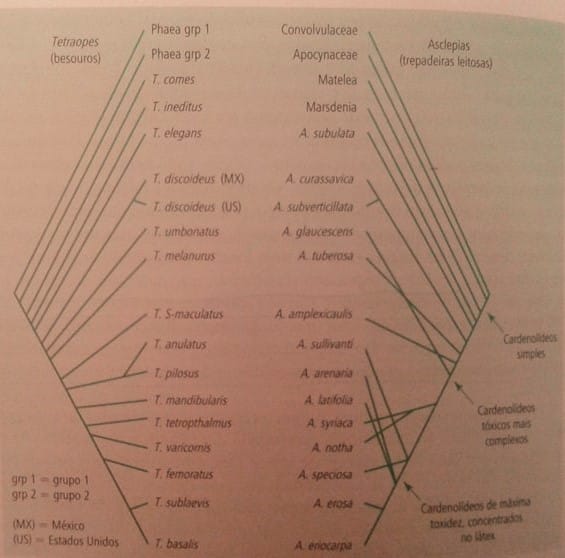 Porém, não necessariamente co-filogenias são resultados de um processo de coevolução, apesar de ser uma forte evidência. A cofilogenia pode surgir também por outros dois processos: evolução sequencial, onde a evolução de uma espécie interfere na evolução da outra, mas o contrário não ocorre e ainda, a especiação de duas espécies que habitam uma região, a partir de uma especiação alopátrica (geográfica) independente da interação das mesmas.
Porém, não necessariamente co-filogenias são resultados de um processo de coevolução, apesar de ser uma forte evidência. A cofilogenia pode surgir também por outros dois processos: evolução sequencial, onde a evolução de uma espécie interfere na evolução da outra, mas o contrário não ocorre e ainda, a especiação de duas espécies que habitam uma região, a partir de uma especiação alopátrica (geográfica) independente da interação das mesmas.
No entanto a cofilogineia não é uma obrigatoriedade para que haja coevolução, por exemplo, algumas plantas produzem substâncias tóxicas que as protegem de possíveis predadores. Os predadores, por sua vez, podem desenvolver defesas contra essas substâncias tóxicas. Isso faz com que, muitas vezes, o inseto troque de hospedeiro e passe a exercer uma pressão seletiva a outras plantas e que, por vez, a planta desenvolva uma nova defesa a partir de novas mutações. Sendo assim, as espécies exercerão influência mútua uma sobre a outra, coevoluindo, porém não apresentarão cofilogenias.
Além disso, a interação recíproca que leva a coevolução pode ocorrer de forma mutualística, onde duas espécies cooperam entre si ou sua interação pode se dar de forma antagonista, ou seja, a partir do conflito/disputa entre espécies. Em última instância, é preciso analisar os casos específicos e os múltiplos aspectos para apontar a ocorrência de coevolução. A seguir analisaremos processos onde fortes evidências indicam a ocorrência de coevolução.
O primeiro caso é a relação entre angiospermas (plantas com flores) e insetos polinizadores. As evidências indicam uma profunda colaboração entre ambos, levando a suas evoluções de forma recíprocas. Os insetos polinizadores e angiospermas se diversificaram simultaneamente durante o período Cretáceo. Hoje o processo de polinização biótico (dependente de outros organismos) é muito mais estabelecido do que a polinização abiótica, assim como uma grande fonte de nutrientes de insetos polinizadores provém de flores das angiospermas indicando sua relação estreita e obrigatória.
Um belo exemplo de coevolução que gerou uma coadaptação entre angiosperma e polinizadores é a fascinante relação entre abelhas euglossinas, conhecidas como abelhas orquídeas e a orquídea do gênero Coryanthes. As abelhas machos não são capazes de sintetizar o feromônio necessário para atrair as fêmeas. Por isso, para produzir o feromônio, o macho coleta inúmeras substâncias de plantas para produzir o cheiro necessário para atrair a fêmea. Para essa abelha, uma das substâncias obrigatórias é produzida pela orquídea do gênero Coryanthes.
Por sua vez, a polinização dessa orquídea ocorre obrigatoriamente com a participação de um processo extremamente encantador. A abelha, ao tentar coletar as substâncias necessárias para produção de seu “perfume”, acaba caindo dentro da flor da orquídea, que é revestida interiormente por uma substância escorregadia que impossibilita que a abelha saia. Porém, no fundo da flor existe um pequeno orifício coberto de pólen. Sendo assim, a abelha só pode sair por esse orifício. Ao sair a abelha sairá com corpo revestido de pólen, tentando coletar mais substâncias para seu feromônio. Em outra orquídea, a abelha poderá cair novamente dentro da flor, a polinizando com o pólen da anterior que acabara de coletar.
 Outro exemplo de coevolução se dá com animais e seus microrganismos benéficos que habitam os organismos. Os seres humanos, por exemplo, apresentam mais bactérias em seu corpo do que células humanas. Parte dessas bactérias são vitais para nossa sobrevivência em inúmeros processos: são fundamentais para a digestão e para sintetizar alguns tipos de vitaminas, protegem nosso corpo contra doenças causadas por bactérias malignas, ou seja, sem elas seríamos incapazes de sobreviver.
Outro exemplo de coevolução se dá com animais e seus microrganismos benéficos que habitam os organismos. Os seres humanos, por exemplo, apresentam mais bactérias em seu corpo do que células humanas. Parte dessas bactérias são vitais para nossa sobrevivência em inúmeros processos: são fundamentais para a digestão e para sintetizar alguns tipos de vitaminas, protegem nosso corpo contra doenças causadas por bactérias malignas, ou seja, sem elas seríamos incapazes de sobreviver.
Por sua vez, essas bactérias precisam de nosso corpo para sobreviver e estão adaptadas a viver em uma série de condições fisiológicas específicas. Nosso corpo e as bactérias que o habitam evoluíram concomitantemente contribuindo para o desenvolvimento específico de cada espécie, mas também produzindo condições evolutivas únicas.
Outro exemplo de coevolução ocorre em situações onde ocorre o mimetismo Mülleriano. Mimetismo é quando uma espécie “imita” outra, mas no caso do mimetismo Mülleriano uns conjuntos de espécies se “imitam”, em geral esse tipo de mimetismo se dá entre espécies não palatáveis (gosto ruim) que evoluem para uma aparência em comum.
Essa é uma estratégia fantástica, na qual a medida que espécies não palatáveis evoluem para uma aparência em comum, o processo de aprendizagem do predador em não se alimentar daquelas espécies é acelerado. O predador precisa aprender apenas uma característica fenotípica, ao invés de várias, assim reduzindo a mortalidade de espécies diferentes que compartilham uma característica em comum. As borboletas vice-rei (Limenitis archippus) e monarca (Danaus plexippus) são impalatáveis e são um caso típico de mimetismo mulleriano.


Todavia, muitas vezes espécies podem desenvolver relações antagônicas, ou seja, os organismos “combatem” um ao outro e, assim, ambos passam a exercer pressões seletivas entre si, fazendo com que passem a coevoluir a medida que uma espécie reponde em relação a adaptação da outra espécie. Um processo clássico é a relação entre parasita e hospedeiro, onde o hospedeiro pode desenvolver adaptações, a fim de combater o parasita e este desenvolve novas adaptações para poder melhor responder as investidas dos hospedeiros.
Uma espécie de coelho (Oryctolagus cuniculus) nativa da Europa foi introduzida na Austrália onde se prosperou de forma muito eficiente, pois na região não existiam seus predadores. A população de coelhos, em questão, apresentou um crescimento populacional desenfreado, se tornando uma praga e destruindo plantações. Para combater os coelhos, em 1950, foi introduzido o vírus mixomatose. Esse vírus tem como hospedeiro natural outra espécie de coelho latino americano.
Em um primeiro momento, o combate aos coelhos com o vírus teve um enorme sucesso, eliminando aproximadamente 99% da população de coelhos em regiões onde o vírus tinha uma alta prevalência. A virulência do mixomatose contra os coelhos era de 100% ,ou seja, todos os coelhos infectados morriam. Porém, a virulência do vírus contra coelho (Oryctolagus cuniculus) passou a sofrer um forte declínio. O decréscimo da virulência foi significativo com o passar dos anos. O declínio da virulência foi demonstrado de forma bastante categórica em uma população de coelhos infectados em laboratório.
Os níveis de virulência foram divididos em 6 graus ( I, II, IIIA, IIIB, IV, V) onde a I é a maior virulência(capacidade de matar) e V a menor virulência. Durante aproximadamente 14 anos de estudos a virulência do mixomatose foi reduzida drasticamente nos coelhos australianos, como demonstra a tabela abaixo:
| Ano | I | II | IIIA | IIIB | IV | V |
| 1950-51 | 100% | 0% | 0% | 0% | 0% | 0% |
| /1958-59 | 0% | 25% | 29% | 27% | 14% | 5% |
| 1963-64 | 0% | 0,3% | 26% | 34% | 31,3% | 8,3% |
Essa mudança da virulência é um exemplo de coevolução, pois a virulência máxima é prejudicial tanto para o parasita como para o hospedeiro. Para o hospedeiro, por razões óbvias, pois caso a virulência seja elevada ele morrerá, porém para um parasita uma virulência elevada também o prejudicará, pois a medida que seu hospedeiro morre precocemente ele terá um decréscimo em seu fitness (w), ou seja, capacidade de deixar descendentes.
A coevolução entre presa e predador é outro caso bastante clássico, muitas vezes, conhecido como corrida armamentista. Esse processo pode ocorrer entre plantas e insetos fitofagos ou entre animais carnívoros e suas presas. Esse processo, basicamente, consiste em uma resposta mútua de defesa e ataque entre presa e predador. A medida que o predador desenvolve melhores mecanismos de predação, uma força evolutiva selecionará as melhores defesas contra esse novo método de ataque que, consequentemente, exercerá uma pressão seletiva para um ataque mais eficiente. Assim, sucessivamente criando uma “escalada evolutiva”.
Um exemplo de corrida armamentista se dá com a lebre e sua predadora, a raposa. A medida que a lebre desenvolveu mecanismos melhores de defesa, como camuflagem ou uma fuga mais eficiente, uma forte pressão seletiva foi exercida sobre as raposas, na qual as que apresentaram melhor resposta adaptativa tiveram melhores chances de sobrevivência e reprodução e que, por sua vez, fizeram com que os genes que desenvolviam essas características que lhe garantissem melhores possibilidades de sobrevivência fossem fixados na população. Por sua vez, agora são as raposas que passam a exercer uma forte força seletiva contra as lebres, que por sua vez terão maior chance de sobrevivência/reprodução, aquelas com mecanismos de defesa, gerando uma resposta mútua de defesa e ataque.
Por fim, podemos concluir que existe uma pressão recíproca entre espécies, onde uma passa a interferir na evolução da outra e vice-versa. Nesse sentido, passamos a ter uma noção mais ampla de evolução, a medida que ela ocorre dentre múltiplas interações, não só com o espaço físico como também com o ambiente biológico entre espécies. Ou seja, a caminhada evolutiva vai além de longa marcha solitária das espécies, mas pelo contrário, é um processo complexo de interação entre as formas de vida que habitam nosso planeta.
Para Saber mais:
- Mark Ridley. Evolução, capítulo 22
- R. Dawkins. O Maior Espetáculo da Terra,capítulo 5,12
- R. Ricklefs. A Economia da Natureza, capítulo 17



