Por Victor Rossetti
Em fevereiro de 2013 um artigo publicado na revista Icarus por matemáticos do Cazaquistão do Astrophysical Institute afirma que o Search for Extraterrestrial Intelligence (SETI) havia encontrado evidências de irredutibilidade e artificialidade no código genético, e que sugeriria uma criação inteligente.
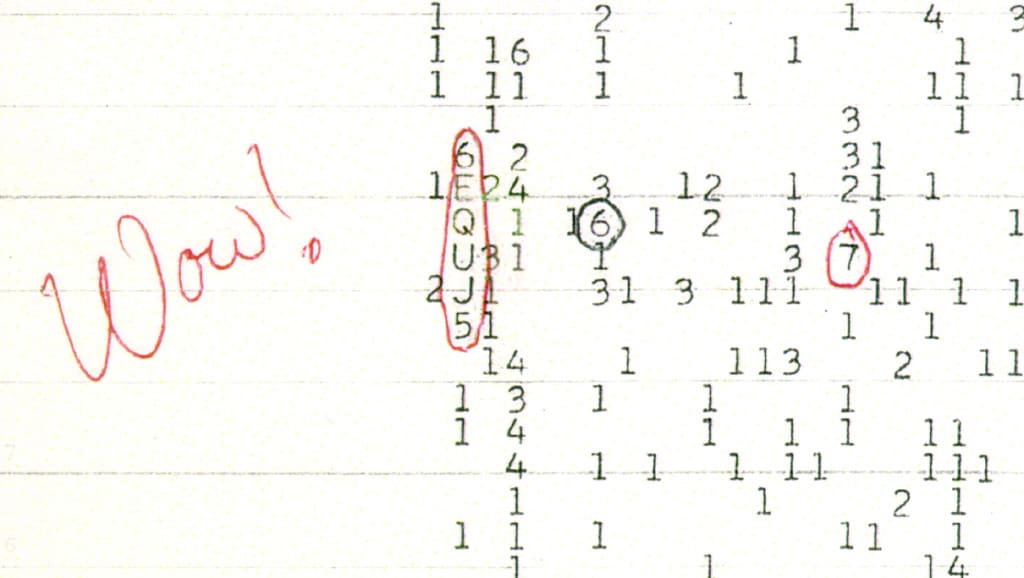
A partir deste artigo é possível fazer algumas reflexões a respeito do que os autores quiseram dizer com código artificial, aspectos matemáticos de sua simetria, seus elementos ideográficos e especialmente a respeito da lista de propriedades estabelecida para evitar falsos positivos e estabelecer se o código genético é realmente resultado de processos inteligentes ou de processos bioquímicos estocásticos (Cherbaka, Makukov, 2013). Sendo assim, pontuo aqui as contradições de tal artigo e como elas desconstroem a ideia de artificialidade do código da vida. O projeto SETI em 1977 recebeu um sinal simples, codificado em letras e números. Os sinais captados pelo SETI geralmente eram bem conhecidos, e se tratavam de emissões radiométricas de galáxias ou outros objetos distantes, sinais quânticos e ruídos de fundo como destaca Carl Sagan em seu livro ”Variedades da Experiência Científica” (2008). Em 1977 um fraco sinal diferente dos demais começou a aumentar gradualmente até atingir seu ápice, decaindo e desaparecendo em seguida. O tempo total de detecção foi de 72 segundos, e seu pico de intensidade era grande o suficiente que ultrapassou o limite de escala de registro.
Concluiu-se que as ondas eletromagnéticas detectadas foram emitidas da constelação de Sagitário, correspondente à linha 21 do hidrogênio. Como o hidrogênio é o elemento mais abundante do Universo a sua frequência natural de emissão é 1420.4556 MHz, e acredita-se que essa também seja a frequência mais óbvia para se tentar algum contato com outras civilizações, tanto para transmissão quanto para recepção de sinais. Como vimos, o sinal “Wow!” foi detectado exatamente nessa frequência.
Sendo assim, se o sinal tivesse vindo de um ponto fixo no espaço, deveria crescer, atingir uma intensidade máxima em algum momento e decair conforme a rotação da Terra movimentasse a antena do SETI. O sinal “Wow!” cumpriu esses pré-requisitos e portanto, foi caracterizado como uma verdadeira emissão vinda de uma fonte fixa do céu de origem desconhecida. A estrela mais próxima que existe na direção de Sagitário está a 220 anos-luz de distância de nossa galáxia. Assim, se o sinal partiu mesmo daquela região, foi um evento astronômico de grande proporção e que até hoje não foi decodificado pelos pesquisadores. Se o sinal demorou 220 anos para chegar a Terra, qualquer tentativa de resposta demoraria mais 220 anos para chegar até o ponto fixo de Sagitário, e se uma civilização inteligente tentou estabelecer algum contato com nós provavelmente já teria desistido de encontrar uma resposta dado o tempo passado desde sua emissão ou mesmo, tal civilização poderia até mesmo ter extinguido (Shuch, 1988).
Desde 1977 nenhum sinal que chamasse a atenção foi novamente detectado. Até agora, mais de 30 anos depois, não se chegou a uma conclusão sobre a origem do sinal, se é que é uma emissão de uma civilização inteligente.
O artigo proposto por esses dois matemáticos do Astrophysical Institute propõem que o DNA é uma molécula artificial. A vida como conhecemos esta baseada em ácidos nucleicos, DNA e proteínas. Estes compostos são responsáveis pela bioquímica da vida. O DNA estoca a informação genética que contém uma receita de como produzir proteínas. Essa informação é transcrita em RNA e traduzida em forma de proteínas, que por sua vez serve como elementos estruturais em tecidos e enzimas (Goldman et al, 2013). Atualmente, o DNA vem sendo usado artificialmente para armazenar informações não-biológicas.
O DNA atua salvaguardando informações sobre como construir um ser vivo. Ele pode ser análogo a informação digital, onde análogo não significa igual, mas sim uma função correspondente. Analogia é uma inferência de significado e não de identidade. O DNA pode ser alterado e ser artificialmente usado para armazenar informações convencionais. Para compreender o que isto quer dizer, mais a frente distinguiremos o significado de sistemas naturais, artificiais e sintéticos. Um estudo publicado na revista Nature, o autor Nick Goldman (2013) armazenou o famoso discurso de Martin Luther King no DNA. Nick deixou claro que apesar de armazenar informações no DNA ele não corresponde a um sistema igual ao de informações gerais. Se de fato fosse idêntico a um sistema biológico as informações artificiais inseridas ali não seriam interpretadas bioquimicamente. Nick Goldman afirma “O DNA que criamos não pode ser incorporado acidentalmente a um genoma, ele usa um código completamente diferente daquele usado em células de organismos vivos… E se esse DNA fosse parar dentro de você, seria degradado e eliminado.”
Como destaca os autores do estudo sobre o “Wow! signal”, o código genético é flexível entre os códons dos aminoácidos e essa flexibilidade permite que um aminoácido tenha 3 ou 4 códons diferentes. Se uma alteração ocorrer no código ela se fixa genoma da espécie, ainda que artificialmente (como nos OGMs e Transgênicos).
Naturalmente, essas variações no genoma são o mecanismo molecular que permite a evolução das espécies (Ridley, 2000). E este conceito darwiniano foi citado, mas não foi considerado no estudo “Wow! signal”, bem como uma série de pressupostos que envolvem o conceito de vida e a biossemiótica.
Uma vez fixado, o código pode ficar inalterado em escalas de tempo cosmológicas. Na verdade, talvez a estrutura mais durável e conhecida na natureza (Cherbaka, Makukov, 2013), especialmente pela sua característica de hereditariedade. Ao afirmar isto, os autores do estudo aceitam tanto a artificialidade do DNA como pressuposto hipotético para a origem da vida, a escala de tempo geológica.
Segundo os autores, o DNA então representa um mecanismo de armazenamento extremamente confiável para se assumir então uma assinatura inteligente, que preenche os requisitos biológicos e termodinâmicos da irredutibilidade de um sistema. Como o cenário atual para a origem da vida na Terra está ainda aberto os autores apresentam a proposta de que a vida poderia ter sido semeada intencionalmente.
De fato, a hipótese ainda esta aberta, mas os autores esbarram em dois problemas grandes; a necessidade de descartar a origem natural (Sagan, 2000); e a diferença entre um sistema de informação artificial de um sistema sintético, apresentando suas respectivas evidências.
Segundo os autores do estudo, um “sinal” inteligente no código genético é estatisticamente testável, portanto, seguiria as normas metodológicas da ciência. Sendo assim, os autores acreditam apresentar arranjos matemáticos simples no código que revelam um conjunto de padrões aritméticos e ideográficos em um único sistema linguístico. Precisos e sistemáticos, esses padrões subjacentes aparecem como um produto de lógica consciente, de precisão e computação trivial ao invés de processos estocásticos como sugere a biologia convencional neo-darwiniana e molecular. Ainda destacam que o sinal apresenta características facilmente reconhecíveis de artificialidade, entre os quais estão; o símbolo zero (na qual a natureza não expressa e portanto é fruto de uma entidade capaz de criar logicamente tal conceito); a sintaxe decimal e semântica simétrica.
Pontuações Conceituais
O que foi notado inicialmente é a ausência de certos conceitos básicos. O que significa o DNA ter origem artificial?
Um sistema natural é obviamente diferente de um sistema artificial. A grande questão conceitual é exatamente a falta de distinção entre um sistema natural, artificial e sintético. Um sistema artificial é um sistema que foi criado pelo homem, mas a partir de manipulação de algo pré-existente. Por exemplo, pesquisadores criaram o primeiro animal com informações artificiais em seu código genético. A técnica poderia dar aos biólogos um controle “átomo por átomo” sobre as moléculas de organismos vivos. Os pesquisadores Sebastian Greiss e Jason Chin (2011) reestruturaram os maquinários biológicos do verme Caenorhabditis elegans para incluir um 21º aminoácido, não encontrado na natureza.
Algo dito sintético é completamente criado pela mão do homem, envolve síntese, ou seja, criação. Uma abordagem evolutiva para a biologia sintética consiste em inocular o processo de evolução por seleção natural em um meio artificial (Ray, 2010).
O código binário e mesmo o decimal podem ser considerados sintéticos. O sistema decimal é uma interpretação matemática que a humanidade criou à alguns milênios. Os registros matemáticos mais antigos datam de 2400 a.C. Progressivamente, o homem foi desenvolvendo sistemas de quantificação e qualificação em seu cotidiano envolvendo medidas de comprimentos e de áreas sugerindo a criação de conceitos. O “Elementos” do grego Euclides (século IV a.C.) foi um dos primeiros livros de matemática que apresentaram de forma sistemática a construção dos teoremas da geometria e foram utilizados no ensino em todo o mundo até ao século XVII (Estrada et al, 2000).
O sistema binário também é sintético. A origem do sistema binário remontam por volta de 3000 a.C. na China que utilizava esse sistema para contagens, somatórias, entre outras operações básicas da matemática. Posteriormente ela foi utilizada na computação. Estes sistemas envolvem números, são sintéticos. Por esta razão, muitas vezes são usados erroneamente para discutir a origem natural do código genético. Porém, um sistema numérico não é passivo de seleção natural e mecanismos biológicos, por se tratar exatamente de números, e portanto é irredutível. O código genético não envolve sistemas numéricos, é flexível para cada aminoácido, e isto é reconhecido até pelos autores do estudo aqui em questão. Fica evidente em:
“For genetic molecular machinery there is no zero, there are nucleotide triplets recognized sterically by release factors at the ribosome. Zero – the supreme abstraction of arithmetic is the interpretive meaning assigned to Stop-codons…”
Esta é uma contradição do artigo, que pode significar um falso positivo e pode custar toda a interpretação do sinal. Outro problema conceitual esta relacionado a definição do emissor de tal sinal.O artigo apresenta uma argumentação que esta relacionada ao conceito de artificialidade, ou seja, de alteração do código genético para estabelecer uma linha de comunicação. Portanto, o artigo deve ser referir ao código como algo alterado e não sintetizado. De fato, ao estabelecer que qualquer alteração de informação se torna fixa no código genético deixa claro que a posição dos autores é tratar da alteração e não criação do código desde suas origens. Como veremos abaixo, os autores não fazem menção alguma sobre a origem do código genético, e sim como eles podem ter sido alterados intencionalmente para emitir uma mensagem.
Alguns proponentes do design inteligente (vertente cristã na qual atribui à Deus o termo de desenhista criador de tudo), defenderam que tal artigo era uma prova científica a favor de Deus, ou um reconhecimento de tal argumento teleológico como científico. Entretanto, o artigo menciona somente alteração no código com uma finalidade específica, comunicação. Sendo assim, a definição mais coerente (se consideramos o sinal real) seria evidência não de assinatura divina, mas de alteração extraterrestre, ou de vida inteligente.
Seja pela interpretação ufológica ou religiosa, o artigo em si não apresenta evidências contundentes a favor da simetria do código. E no decorrer deste texto decifraremos não o código, mas a metodologia matemática que nos conduz ao código e suas respectivas abstrações.
Os autores levantam uma lista de características que devem ser consideradas para estabelecer se o código genético é resultado de processos artificiais e como evitar falsos positivos.
Para ser considerado, de forma inequívoca, um sinal inteligente, os autores consideraram que o sinal deve ser altamente significativo estatisticamente e não só devem possuir características semelhantes à de inteligência, como deve ser incompatível com qualquer processo natural, seja a evolução darwiniana ou lamarckiana, não deve ser impulsionado pela biossíntese de aminoácidos, nem por alterações genômicas ou por afinidades entre anti-códons e aminoácidos. Além disto, não pode ser compatível com processos de seleção que levam ao aumento da diversidade de proteínas energéticas das interações códon/anticódon, ou vários mecanismos pré- translacionais (Cherbaka, Makukov, 2013).
Para não ser considerado um falso positivo no que diz respeito a interpretação do sinal, os símbolos devem ser matemáticos, presumindo que a linguagem numérica é a mesma em todo o universo, tais como sistemas numerais posicionais, como o conceito de zero que não ocorre na natureza e portanto seria uma característica de inteligência. Assim, as mensagens interestelares enviadas da Terra geralmente começam com sequência natural de números em notação binária ou decimal. Ou seja, ele deve seguir um sistema numérico criado por nós e presumidamente universal, mesmo que este código seja sintético, criado pelo homem.
Para reforçar a artificialidade, essas mensagens também devem incluir símbolos de operações aritméticas, triângulo egípcio, DNA e outras noções de consciência humana.
Entretanto, sabemos que o DNA, que é um componente químico e natural não tem qualquer significação ligada ao zero (especialmente bioquímica), portanto, ou o resultado seria nulo, ou um falso positivo adquirido no uso de elementos numéricos para estabelecer um conceito numeral como o zero, atribuído a uma base do DNA. Sendo assim, o reconhecimento do zero no código poderia ser simplesmente resultado de nosso sistema aritmético.
Em segundo lugar, para minimizar o risco de falsos positivos necessita-se de requisitos que sejam o mais restritivo possível em um sinal putativo. Por exemplo, é razoável esperar que uma mensagem genuinamente inteligente não ira representar apenas uma coleção de padrões de vários tipos, mas os padrões do mesmo “estilo linguístico”, neste caso, um sistema matemático simétrico. Outra exigência rigorosa pode ser que os padrões devam envolver cada um dos elementos do arranjo do código, enquanto que todo o sinal deve ocupar a maior parte, se não todos, de capacidade informativa do código. Sendo assim, todo o sistema de códons e seus respectivos aminoácidos devem seguir o mesmo sistema numérico. E de fato, seguem na decomposição feita pelo autor, mas como resultado de uma matemática simples, baseada na razão 3 como veremos.
O maior viés de todos é que ele esbarra em algo que os autores não tocaram. A origem das bases nucleicas e do código genético por um mecanismo natural. De fato, isto é abandonado pelos autores mesmo que a literatura atual esteja bastante embasada (Cherbaka, Makukov, 2013).
Para estabelecer um paradigma como vigente, é preciso que a hipótese consiga se mostrar epistemologicamente mais promissora que as concorrentes ou a vigente (Mendonça, 2002). Os autores não discutem qualquer hipótese ou teoria sobre a origem dos nucleotídeos e do código genético. Ao rejeitar as explicações a partir de mecanismos naturais os autores partem desde o principio que a vida inteligente existe, e não que o código genético possa vir conter um código que sugira criação inteligente. Portanto, a pesquisa inicial do autor não é baseada em hipóteses que serão avaliadas ao decurso e ao final do artigo. Ao contrário, os autores iniciam o estudo com uma ideia pré-concebida de tal forma que a matemática vai de encontro a simetria aritmética dentro do código genético. Este pode ser um grande falso positivo, pois a pesquisa pode não ter sido conduzida livremente, mas sim direcionada, e o trecho do artigo em que isto fica explicito é:
“We show that the signal is statistically significant, employs informational capacity of the code entirely, and is untraceable to natural origin. The models of emergence of primordial life with original signal-free genetic code are beyond the scope of this paper.”
Em síntese, o estudo se baseou em uma interpretação da matemática, uma ciência exata enquanto o código genético faz parte de um conceito respectivo as ciências biológicas, que é uma ciência não exata e sim natural. O estudo pode sim seguir em direção a um falso positivo quando atributos absolutos preponderados pela matemática tomam partido em um sistema natural.
Origem do DNA e Evolução do Código Genético
Antes de discutir a possibilidade do DNA e códon terem surgido por processos artificiais, faz-se fundamental discutir se tais processos biológicos não são primeiramente fruto de processos naturais. Em ciência, a resposta mais simples geralmente é a correta, pois respeita a navalha de Ockham.
Kuzicheva e SImakov (1999) em seus experimentos indicam que nucleotídeos, os blocos de construção primários do DNA, poderiam ser sintetizados em um estágio inicial da evolução do Sistema Solar e transportados em segurança para a Terra por cometas, asteróides e partículas de poeira. Outros estudos apontam que em meio argiloso (Luo et al, 2013), ou aquoso (em superfície oceânica), é possível que o DNA tenha surgido e se ligado de tal forma a permitir a origem da vida (Michaelian, 2011). Simulações feitas na água do mar mostram que a argila forma um hidrogel, uma massa de espaços microscópicos capazes de absorver líquidos como uma esponja. Ao longo de bilhões de anos, os produtos químicos confinados nesses espaços poderiam ter realizado as reações complexas que formaram as proteínas, DNA e, eventualmente, todo o maquinário que faz um trabalho de célula viva. Hidrogéis de argila poderiam ter esses processos químicos confinados e protegidos até que a membrana que rodeia as células vivas surgisse (Luo et al, 2013). Os estudos modernos favorecem o fato de que as moléculas orgânicas extraterrestres serviram como uma importante fonte de produtos químicos biológicos importantes na Terra primitiva. Presume-se que estas moléculas orgânicas feitas no espaço possam ter sido transportadas com segurança para a superfície da Terra associadas a grãos minerais.
Tem sido mostrado, que no vácuo o ultravioleta (145 nm) promove a síntese de nucleotídeos de forma mais eficaz do que o ultravioleta (UV 254 nm). A formação de 5′-mononucleótidos parece ser a reação mais eficaz tanto em voo de meteoros como já foi demonstrado empiricamente quanto em experiências de laboratório (Kuzicheva e SImakov, 2002).
A Terra recebe 40 mil toneladas de material proveniente de cometas, alguns dos quais são condrito carbonáceo, que estão entre os materiais mais antigos do Sistema Solar com elevada concentração de carbono (cerca de 3%). Um dos mais estudados meteoritos deste tipo caiu na Austrália em 1969 e recebeu o nome da localidade, Murchinson. Análises deste meteorito demonstraram a presença de 500 compostos orgânicos e 80 aminoácidos (Kwok, 2009).
De fato, até compostos como o ácido pirúvico, ácido oxaloacético, ácido cítrico, ácido isocítrico, e ácido cetoglutárico-α (todos os membros do ciclo de ácido cítrico) não foram identificados em fontes extraterrestres, mas em meteoritos carbonáceos (Cooper et al, 2011). Deste então, a análise de outros meteoritos deste tipo tem demonstrado características semelhantes. Mais de 140 moléculas orgânicas foram encontradas no espaço (Kwok, 2009).
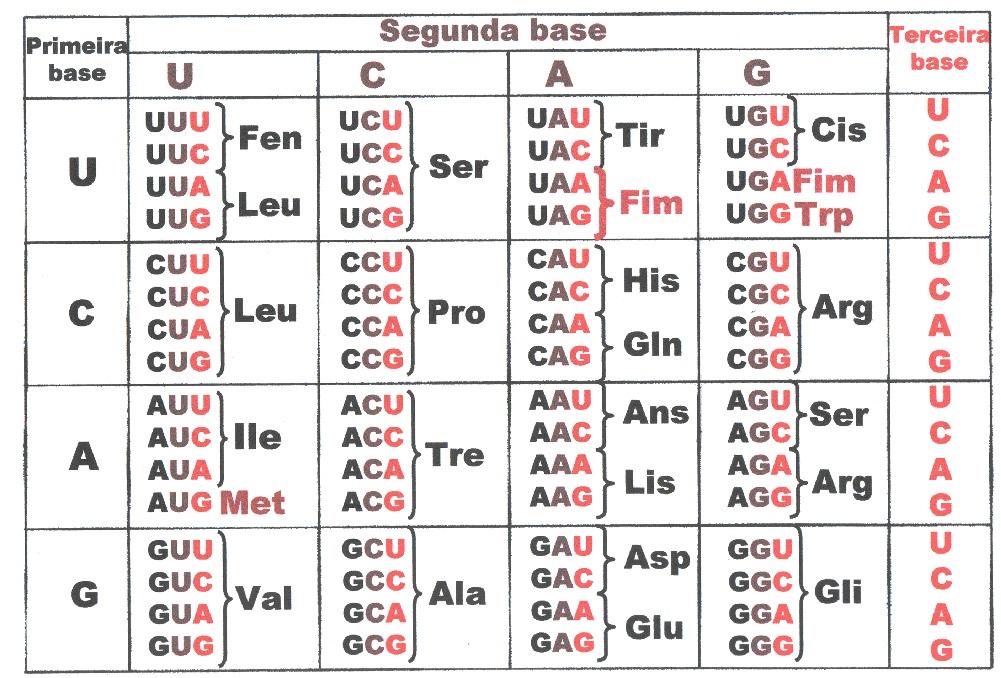
Quando os primeiros blocos nucleicos de replicação se consolidaram como responsáveis pela hereditariedade (seja o DNA, ou no RNA World ou em moléculas anteriores) a replicação e manutenção do metabolismo começou a depender de produtos que seriam sintetizados pelo DNA ou RNA e seu código genético, além de elementos químicos disponíveis no ambiente. Estabeleceu-se então o código genético, que é baseado em um sistema de códon (ou triplex), onde cada trinca corresponde a um aminoácido da proteína. É fundamental entender este fenômeno, pois a trinca é a grande chave para compreender a interpretação do sinal, e a simetria que os autores encontraram no DNA. (Veja o vídeo).
A questão é; porque um códon é formado por três nucleotídeos, e não um ou dois?
A trinca se estabeleceu por mecanismos químicos de otimização e redução de erros tornando o código estável com 3 nucleotídeos. Há um conjunto de teorias que são complementares entre si e que podem explicar a origem de tais códons sob o ponto de vista natural. A teoria da coevolução do código genético de Wong (2005) e vias biossintéticas de aminoácidos na sua versão atual pode explicar o fato de que alguns dos aminoácidos ligados biossinteticamente têm a mesma base na primeira posição.
De acordo com esta teoria a codificação de aminoácidos ocorreu sequencialmente. Eles foram sendo estabelecidos e amadurecendo sequencialmente suas respectivas vias biossintéticas de aminoácidos com reações de intervenção energeticamente favoráveis, atribuindo códons em diferentes nucleotídeos. Isso pode ter ocorrido por muitos motivos; porque eles são quimicamente semelhantes; pela interação entre aminoácidos e a tríade dos códons; pela interação de um novo aminoácido ou a interação de um intermediário metabólico deste sítio de ligação com o aminoácido substituído na enzima aminoacil-RNAt sintetase.
Isso tem respaldo empírico, uma vez que verificou-se os aminoácidos incluídos na mesma família biossintética, como o aspartato (GAU e GAC) ou glutamato (GAA e GAG), têm a mesma base na primeira posição códon. Isso evidencia códons mais primitivos.
A teoria estereoquímica explica a associação entre a natureza da base que está na segunda posição e a do aminoácido. Em muitos casos, os mesmos aminoácidos são designados por códons que diferem entre si apenas na natureza da terceira base por que há semelhanças em certas propriedades físico-químicas atribuídas aos códons vizinhos com a mesma base na segunda posição (Ckrick, 1968).
De acordo com esta teoria, os padrões observados no código genético ocorrem devido à interação entre os aminoácidos e códons próprios, anticódons e códon. Essa interação ocorre por requisitos polares que não é mais do que o coeficiente de partição de aminoácidos no sistema de água-piridina. Ele varia de acordo com a natureza da base da segunda posição. Há também a correlação entre os nucleotídeos e a hidrofobicidade da primeira e segunda posição do códon e até mesmo entre as forças de Van der Waals dos grupos amina que estão livres e interagem o hidrogênio do códon com o anticódon.
Em todos estes casos (exceto o último), foram obtidos provas de uma interação direta entre o aminoácido e RNA. Em laboratório, experiências de amplificação e seleção in vitro isolaram sítios de RNA com ligação a vários aminoácidos como a arginina, valina e isoleucina. Acredita-se que no RNA–World de alguns bilhões de anos atrás os aminoácidos eram ligados entre si por interações covalentes e não por ribozimas primitivas. As ribozimas que catalisavam reações de pequenos peptídios passaram a sintetizar diferentes complexos proteicos de RNAt e RNAm muito semelhante ao aparato enzimático existente atualmente na tradução (Lazcano, 2007).
Assim, a interação de um aminoácido com o códon de RNA é muito semelhante ao que existe no códon vizinho. Isso é atribuído em função da lei de minimização de erros no código genético que é uma lei puramente química e não uma variação aleatória ou sobre a atuação da seleção natural. É uma propriedade química natural. Essas propriedades estruturais do código são explicadas dentro da teoria da otimização do código genético, ou de minimização de erros.
Isso quer dizer que existe uma espécie de filtro de erros já que eles ocorrem nos processos básicos de replicação, transcrição e tradução. Quando isso afeta a função básica de reconhecimento do significado estabelecido desses códons pode custar a sobrevivência do organismo.
Se notarmos, o quadro de aminoácidos e seus respectivos códons, veremos que as primeiras e a segundas bases são idênticas. Isso quer dizer que aminoácidos como a alanina sempre vão começar com as bases GC. Mais tarde, Carl Woese (1977) propôs a teoria de erros de tradução já que nas fases iniciais de decodificação do códon as ligações químicas eram muito instáveis e ineficientes.
Muitos erros de tradução estão envolvidos na leitura do códon em relação ao códon vizinho. Para refinar ainda mais a relação genótipo/fenótipo/ambiente a evolução favoreceu a atribuição mais precisa de aminoácidos com códons semelhantes. Desde o trabalho inicial deste tipo até hoje foram desenvolvidos diversos métodos para quantificar o grau de otimização do código genético em relação aos códigos aleatórios, especialmente a obtenção de amostras aleatórias de códigos até características da hidrofobicidade, exigências polares, ponto isoelétrico, tamanho molecular e refratividade dos aminoácidos e códons.
Todas essas teorias são perfeitamente compatíveis entre si e explicam a conformação atual do código genético que é universal, e portanto, fruto de processos evolutivos também (Michaelian, 2011). Após a conformação do código a partir de processos evolutivos a vida se diversificou partindo do ancestral comum a toda vida (chamado de LUCA ou Last Universal Common Ancestor) dando a conformação sistemática que conhecemos hoje (Lazcano, 2007, 2008).
Desconstrução Matemática da Hipótese Simétrica e Ideográfica
Sabendo das propostas da ciência sobre a origem dos blocos de construção e do DNA é possível entender a lógica matemática usada pelos autores no artigo para estabelecer uma simetria aritmética.
A simetria é a chave de todo o sinal. Se conceitualmente, os autores deixaram em aberto questões básicas e práticas, sendo uma abordagem matemática, ou seja, exata, qualquer quebra simétrica descaracteriza todo o sinal, incluindo sua interpretação ideográfica. Tal simetria foi obtida a partir dos códons. O códon é formado por três bases e corresponde a um aminoácido da proteína. Essa relação 3 nucleotídeos/aminoácido é a razão matemática na qual o sinal foi interpretado. Isto quer dizer que ele foi construído segundo uma interpretação matemática que é lógica, exata, e portanto é meramente resultado da decomposição de números com base na razão 3.
O artigo apresenta a simetria pelo número 037. É através dele que toda simetria do código surge a partir do sistema decimal. Simetria então, significa que se usarmos uma progressão aritmética de base 037 e colocarmos cada algarismo de sua composição no lugar de um nucleotídeo de cada códon há uma simetria numérica perfeita se forem dispostas em 3 fileiras de 9 códons cada. Se o código com base 3, obviamente o número 37 ganha um a esquerda para que assim preencha os requisitos da razão utilizada. Porém, este conceito de zero foi introduzido pelas autores e não corresponde simetricamente e bioquimicamente ao códon, o que caracteriza outro viés.
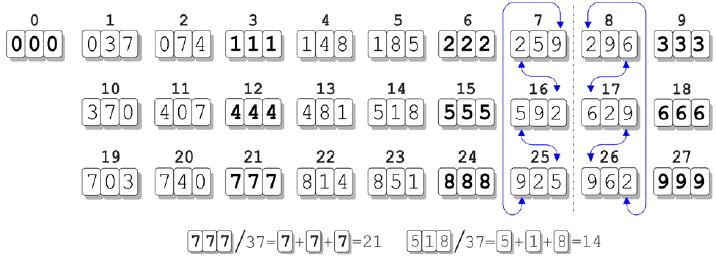
Apesar de ser uma progressão aritmética, na qual decorre a partir do número 37, a explicação para tal fenômeno de repetição de números simétricos nas colunas é resultado do produto de números múltiplos. Isso decorre da razão 3.
O produto do número 37 pelos primeiros múltiplos de 3 é sempre esta sequencia simétrica; 111, 222, 333, 444… até 999 que é limitado pelas 3 casas do códon. Acima disto, a simetria é quebrada, ou seja, a simetria deixa de existir se o produto passar de 3 casas decimais na razão 3. Assim seria se a razão fosse 5 e o código genético fosse estabelecido com 5 nucleotídeos.
O maior divisor primo dos números 222, 333, 444 e 555 é portanto 37, e ao realizar uma progressão aritmética estes números aparecem.
Visualizando por outro ângulo para tirar a prova dela, temos a razão 3 e quando a progressão é feita até o último número de 3 algarismos (que é 999) e logicamente 9+9+9 = 27 (ou 3 linhas x 9 colunas = 27), que é divisível por três. Logo 999 é o último termo com 3 dígitos de uma progressão aritmética onde a razão é 3. Se dividirmos 999 por 3 (que é o número de algarismo no códon) teremos 333. Trezentos e trinta e três é divisível por 3 cujo produto é 111. Se dividirmos este número por 3 novamente teremos o número 37.
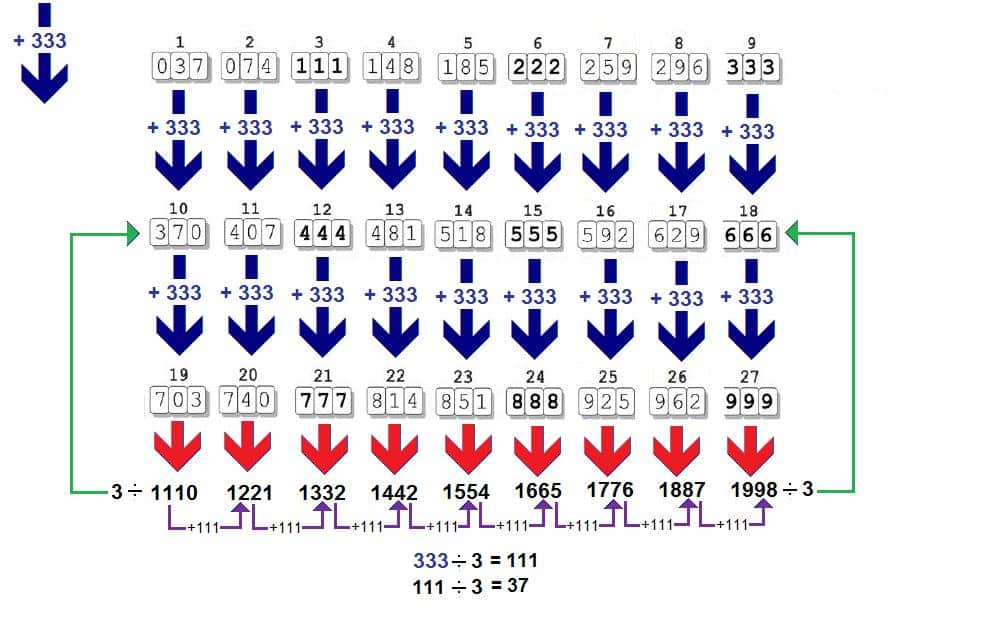
A soma de 333 entre os elementos da coluna sempre vai gerar um produto divisível 3 por que ele é o número médio de cada coluna. Se dividirmos esses produtos sempre por três teremos no final o número 37. (1.110/3 = 370, ou seja, o segundo valor da coluna 1, e assim com as demais colunas).
O número 37 apresentado no artigo, nada mais é do que uma brincadeira feita por matemáticos à décadas, usando a razão 3 ou o 37 que matematicamente é apelidado de “número mágico”.
O número três é matematicamente flexível e está envolvido em muitos outros princípios matemáticos, e coincidentemente, até mesmo biológicos. Por exemplo, o número três é o primeiro dos números Primos de Fermat. Na matemática, o número de Fermat é um número inteiro positivo que assume a forma segundo a fórmula Fn= (2)2n+1 onde n um número natural. O algarismo 3 também é o primeiro número primo de Mersenne.
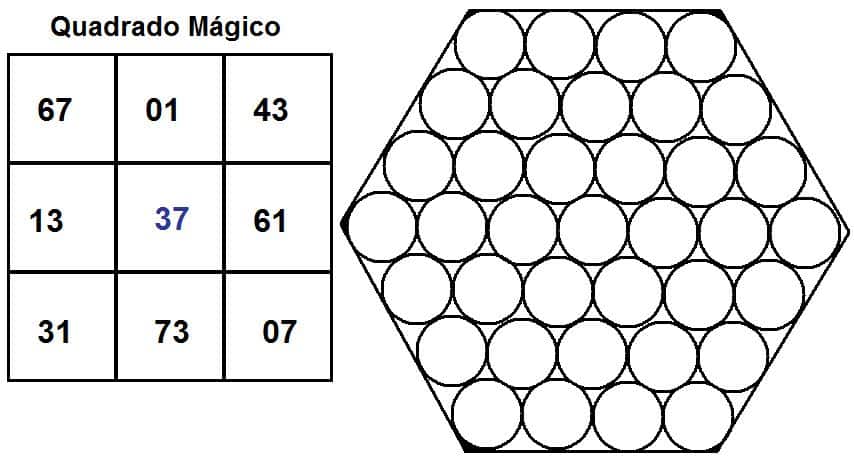
O algarismo 37 também é um número primo e também tem propriedades geométricas, pois é o número hexagonal centrado. Na matemática, ele é o numero chave do “quadrado mágico” cuja soma das linhas é sempre 111, ou seja, um múltiplo de 3, cujo produto é 37. Na biologia ainda temos o número 37 identificando a quantidade de genes mitocondriais, ou mesmo, a quantidade de músculos envolvidos na movimentação da mão.
Entretanto, o suposto sinal não acaba neste ponto. Segundo os autores, cada aminoácido apresenta um mesmo número, específico, que é o 74. Este representa o número atômico de uma parte específica de cada aminoácido. São dos átomos que pertencem somente aos principais pontos de ligação entre os aminoácidos, os sítios aminos e carboxílicos. O que os matemáticos fizeram foi selecionar o grupo amino e carboxílicos de cada aminoácido e calcularam o número atômico deles, onde C2(2×12),N1(1×14),O2(2×16), H4(4×1) como pode ser visto no aminoácido abaixo:
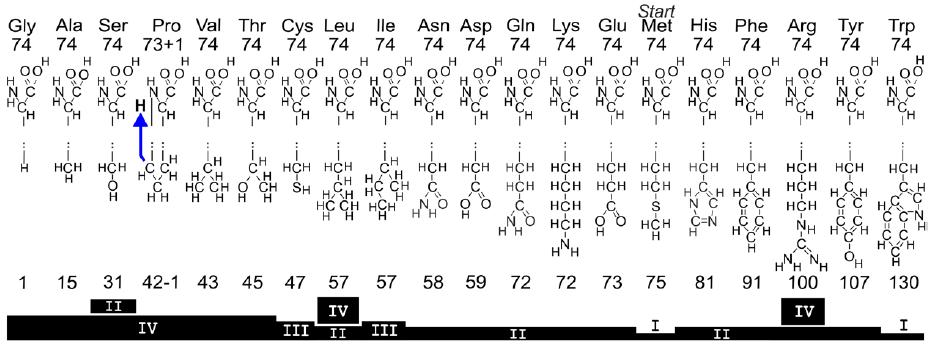
Os autores rejeitaram o radical dos aminoácidos porque alteraria o número atômico. Em suma, o número 74 é resultado de um cálculo comum a quase todos os aminoácidos, o que é óbvio, considerando que o que caracteriza o aminoácido é justamente a presença de uma extremidade amina (número atômico 17) e outra carboxila (número atômico 57). Por consequência, todos os aminoácidos não coincidentemente tem o mesmo número de átomos (74). A prolina é a única exceção, pois carrega um hidrogênio a menos e portanto, não codifica sinal algum e não foi considerada um viés no estudo. Ela foi simplesmente descartada, o que caracteriza um viés uma vez que uma das condições pontuadas pelos autores para que não seja considerado um falso positivo é “…should be inconsistent in principle with… driven by amino acid biosynthesis, genomic changes, affinities between (anti)codons and amino acids, selection for the increased diversity of proteins energetics of codon-anticodon interactions, or various pre-translational mechanisms” como já foi mencionado anteriormente. Note que o autor simplesmente desconhece, ou rejeita qualquer explicação natural para a prolina ter uma cadeia lateral, e um hidrogênio a menos: “Therefore, there is no any natural reason for nucleon transfer in proline; it can be simulated only in the mind of a recipient to achieve the array of amino acids with uniform structure. Such nucleon transfer thus appears artificial.” Usando um sofisma, ou silogismo dicotômico em que se A não explica a origem de B, portanto C explica.
As aminas são compostos químicos orgânicos nitrogenados derivados do amoníaco (NH3) e resultam da substituição parcial ou total dos hidrogênios da molécula por grupos hidrocarbônicos (ou seja radicais alquilo ou arilo abreviados pela letra R). Se substituirmos um, dois ou três átomos de hidrogênio, teremos, respectivamente, aminas primárias (R-NH2), secundárias(R1R2NH) ou terciárias (R1R2R3N). Portanto, é natural que a prolina possa ter um hidrogênio a menos, e sua estrutura anelar resulta da ligação do terminal alfa-amina (NH2) à cadeia variável alifática.
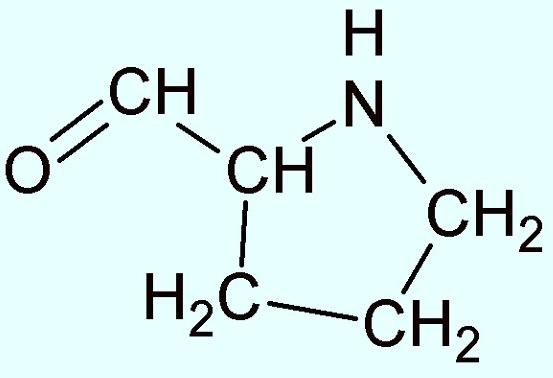
Uma razão para que o nitrogênio tenha uma ligação a menos é que este elemento possui 5 elétrons em sua camada de valência e para chegar a sua estabilidade precisa de mais 3 elétrons, por meio de 3 ligações, que são geralmente feitas com carbono, hidrogênio ou oxigênio. Então, o nitrogênio faz somente 3 ligações químicas, por essa razão é mais comum encontra-lo em NH2 ligado a um carbono.
Os autores juntam a simetria do códon de base 37 com o 74 a partir dessas duas extremidades dos aminoácidos com uma simples multiplicação. A conexão é simples, eles multiplicam o número 37 pelo número de extremidades nos aminoácidos (amino e carboxila) que é 2 e obtém o 74. Eles coincidem o fato de o número atômico ser 74 com o fato do 37 ser um número simétrico no códon multiplicado pelas duas extremidades. Ao multiplicar criam uma suposta ligação matemática, simétrica, aritmética e ideográfica com base nas extremidades do aminoácido, mesmo que elas não tenham números atômicos proporcionais. O número atômico das aminas é 17 enquanto o carboxílico é 57.
Discussão
O estudo apresentado acima oferece somente uma suposta simetria, criada a partir de interpretações numéricas de acordo com a forma com que o código genético evoluiu, portanto, é fruto de processos naturais como abordado acima, sobre a perspectiva da química. A otimização e redução de erros de síntese proteica foram fatores fundamentais para a caracterização atual do código genético, que é universal em todos o grandes domínios da biologia (Arquea, Bactéria e Eucaria) e portanto evidência a origem monofilética desses grupos (Lazcano, 2007, 2008). A simetria de inteligência é aparente, fruto de um sistema numérico humano em uma reinterpretação algébrica. Como ressaltaram os autores, a natureza não interpreta o número zero, e o sistema utilizado pelos autores para traduzir o suposto sinal inteligente é numérico. Portanto, o sinal interpretado no estudo foi identificado como sinal de inteligência, e de fato é, um sinal criado por nós mesmos em nosso sistema decimal. A decodificação foi projetada e identificada por nós, porém projetada a uma inteligência exógena. O numero 37 é uma projeção de nossa matemática associado a razão 3 e portanto é um sinal de raciocínio lógico-matemático. O DNA é um sistema de armazenamento de informação natural, e não interpreta números ou algarismos binários. Por mais que sinteticamente tenhamos criados sistemas de informações análogas, eles somente têm a função igual, não o modo de armazenamento igual, caso contrário Nick Goldman conseguiria interpretar informações convencionais bioquimicamente, em proteínas.
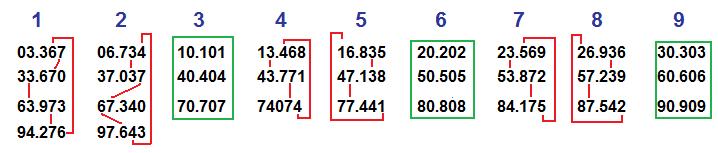
Um sistema que trabalhe com 5 algarismos e que inicie com o número 03.367 também poderia apresentar o mesmo tipo de simetria que o apresentado no trabalho desses matemáticos. Porém, assumidamente sintético.
Ao olhar um sistema biológico, é natural atribuir finalidades e valores estéticos quando se observa em uma escala de tempo relativamente curta. Os animais que vemos hoje nos ecossistemas, preenchendo um determinado nicho ecológico, com seu elenco de genes e suas características físicas, embrionárias, comportamentais e estéticas (Primack & Rodrigues, 2001) é fruto de processos evolutivos como descrito por Charles Darwin (1859) em seu livro “A Origem das Espécies”. A evolução das espécies é um processo dinâmico, cumulativo que pune com a morte os inaptos, favorecendo a sobrevivência daqueles que são frutos de uma descendência com modificações. Este processo não é notado em tempo real, pois a seleção natural e tantos outros mecanismos evolutivos (seleção sexual, deriva genética, epigenética, equilíbrio pontuado, exaptação…) trabalham através de gerações. Por essa razão, atribuímos que um determinado animal está perfeitamente adaptado a um determinado ambiente e parece ter sido criado para desempenhar uma determinada atribuição ecológica. O que vemos é sempre o estado final de um processo natural que se iniciou a cerca de 3,5 bilhões de anos. Este resultado não é final, e sim um estado, até que o animal consiga se reproduzir e passar seu diferencial a próxima geração, Assim, o que vemos é um resultado aparente e não conclusivo, diante da dinâmica que é o processo evolutivo. Neste contexto todo, podemos ver a evolução sob duas óticas distintas, na qual o zoólogo Richard Dawkins (1973) expõem em seu livro o “Gene Egoísta”. A seleção atuando no nível das espécies, punindo com a morte os inaptos; e a evolução sob o ponto de vista dos genes, como eles cooperam entre si favorecendo o alcance a próxima geração. Os genes (+/- 25 mil) são porções do DNA que contém as informações para sintetizar proteínas (+/- 80 mil), definir aspectos metabólicos, fisiológicos e comportamentais das diferentes formas de vida.
Embora o conceito de vida ainda esteja aberto dentro do campo da filosofia da ciência, em especial da filosofia da biologia, listas de propriedades que caracterizam biologicamente um ser vivo foram criadas e abarcam 3 conceitos de vida igualmente importantes e vigentes, baseados em replicadores darwinianos, autopoiese e a biossemiótica (Correa et al, 2008).
Os autores do presente artigo tocam no caso da biossemiótica. Porém, como ressalta El-Hani (2000), a produção, emissão e interpretação de signos aparentemente não surgiram apenas na espécie humana, mas estaria aqui desde a origem da vida. Para a biossemiótica, a natureza é cheia de signos e é parte desta disciplina tentar reconstruir a história natural desses signos, descrevendo a evolução de diferentes sistemas de signos, de interpretação, seja ele um sistema genético ou mesmo a linguagem humana. Alguns pesquisadores e filósofos criticam a ideia de usar o conceito de informação na biologia, uma vez que não há uma noção concreta do que significa informação. Desta forma, a informação genética seria uma metáfora utilizada na biologia que poderia conduzir a uma visão errônea da explicação em campos como a biologia molecular (Stuart, 1985 apud El-Hani, & Videira, 2000). Os biossemioticistas discordam dessa posição, afirmando que o uso de conceitos e teorias da semiótica na interpretação da “linguagem” da informação biológica contribuiria exatamente para sua formulação de maneira precisa e consistente e até mesmo didática para a compreensão do papel que o código genético tem na biologia dos seres vivos.
No artigo, os autores sustentam a ideia de que uma maneira de encontrar padrões irredutíveis a eventos naturais é envolvendo semântica, já que nenhum processo natural é capaz de interpretar símbolos abstratos. Isso contraria com a biologia uma vez que a natureza é formada por signos, ou elementos que podem gerar significados e faz parte da biossemiótica reconstruir a história desses signos, descrevendo sua evolução em diferentes sistemas.
Sendo assim, os autores prosseguem afirmando que o código genético em si é considerado como uma “convenção natural” e referem-se aos códons como símbolos e aos aminoácidos como seus significados. De fato, o código genético pode ser visto como um sistema de signos que representa conjuntos de reações químicas através de um sistema formal de interpretação na biologia.
Para os autores, estas abordagens fazem distinção entre semântica orgânica de códigos moleculares e semântica interpretativas ou linguísticas peculiares a inteligência. E alegam que este último tipo de semântica é revelada no sinal do código genético. É apresentada não só na simetria de sinais sintáticos antônimos, como também no símbolo de zero. Para maquinaria molecular genética não existe zero, há triplex de nucleotídeos reconhecidos estericamente por fatores de liberação no ribossomo. Assim, vemos que o conceito lógico de zero é a abstração suprema da aritmética, é o significado interpretativo atribuído ao códon-Stop, ou códons de parada (UAA, UAG e UGA).
Entretanto, os códons de parada, que representam o zero na linguagem decimal utilizada pelos autores é quimicamente muito mais aprimorado do que simplesmente o significado de zero. Ele contém eventos bioquímicos complexos. O término de tradução proteica ocorre quando um códon de parada ocorre no sítio A da subunidade 60S do ribossomo. O fator de terminação eRF, se liga a uma molécula GTP, que estimula a ribozima peptidil-transferase para transferir o péptideo do RNAt no sitio P, juntamente com a hidrólise de GTP em GDP + PI. A proteína é então liberada a partir do ribossomo ao longo do complexo RNAt/eRF/GDP. Os fatores anti-associação em seguida promove a dissociação das duas subunidades ribossomais para que uma nova síntese proteica possa ocorrer (King, 2013).
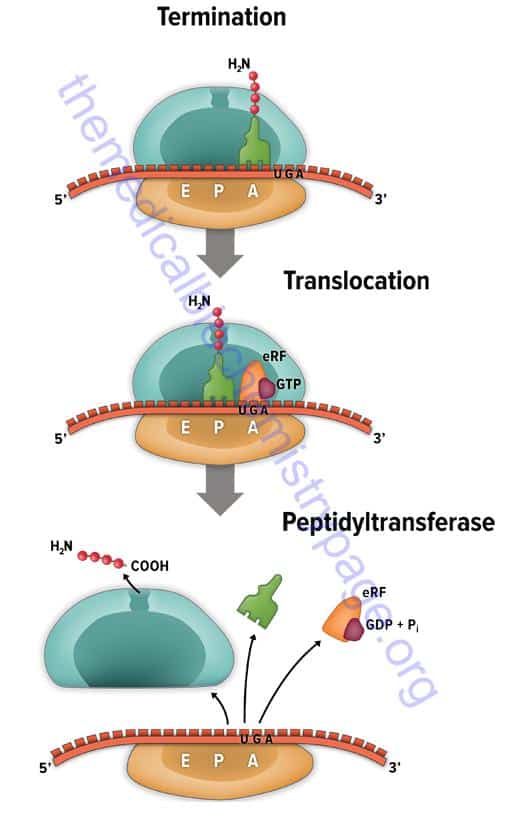
No que diz respeito a possibilidade do artigo fomentar algum tipo de design inteligente, fica evidente pela própria descrição do artigo, que se de fato o DNA é resultado de processos artificiais, ele é fruto de alteração e não de síntese total. Faz-se fundamental distinguir a criação da manipulação de um códon. A matemática apresentado pelos autores é simples, mesmo quando introduzida dentro de um código de razão 3, mas poderia justificar irredutibilidade de sistemas somente dentro dos parâmetros matemáticos, e portanto, dentro de nossa linguagem, não refletindo os valores reais dos eventos, mas nossos valores numéricos. Isso ficou evidente ao demonstrar que nucleotídeos podem surgir na Terra ou no espaço de forma plenamente natural; ou que o códon de parada obtém um significa zero quando atribuímos valores numéricos a ele. Em instância real, nos processos bioquímicos, um códon de parada gera uma série de reações subsequências envolvendo maquinários enzimáticos e hidrolíticos que estão além da mera interpretação nula apresentada pelos autores. Qualificar em números processos biológicos e químicos complexos pode fomentar falsos positivos, e justificar alegações pseudocientíficas, como é o caso do design inteligente. É praxi de tal grupo utilizar conceitos como complexidade irredutível, complexidade especificada ou mesmo algoritmos matemáticos que justifiquem crenças teleológicas pessoais. Talvez o caso mais comum que demonstre como estes falsos positivos surgem a partir de matemáticas tendenciosas seja a extrapolação da Lei de Borel.
A probabilidade de um evento é expressa como um número real de 0 a 1, sendo mais provável quanto maior for o número. Um evento pode ter apenas uma probabilidade, a qualquer momento, No entanto, uma forma extrapolada da Lei de Borel usada por proponentes do design inteligente é implicar em absurdos matemáticos, onde eventos de baixa probabilidade ganham duas atribuições probabilísticas diferentes; sua verdadeira probabilidade e uma probabilidade de 0.
Se um computador gerar 100 dígitos aleatórios, haveria 10100 resultados igualmente prováveis e possíveis. A probabilidade de um determinado resultado seria, assim, 10-100. Aplicando a “Lei do Acaso” segundo os defensores do design inteligente, teríamos de concluir que qualquer resultado é concebível, porque ele tem uma probabilidade de menos de 1 em 1050. Portanto, literalmente impossível, não tendo possibilidade de ocorrência, assim, teria uma probabilidade de 0. É lógico que nenhum evento pode ter uma probabilidade de 1 em 10100 e ter probabilidade de 0. Além disso, uma vez que os resultados possíveis são o que os matemáticos chamam mutuamente exclusivos e exaustivos em conjunto, a soma de todas as suas probabilidades individuais deve ser igual a 1, o que eles não podem fazer se todos forem 0 (Robson, 2000).
A ideia de encontrar sinais aritméticos simples pode indicar viés em um sistema, que pode ser simples o suficiente para ser encontrado em coisas não inteligentes. Porém, se real, ser codificável pelo homem. Portanto, como saber se o código tem sinais de inteligência ou se é fruto de nossa própria abstração? O número zero pode se encaixar nessa contradição.
Sinais poderiam ser considerados fruto de inteligência se houvesse padrões estatísticos consideráveis que indiquem a decomposição de cálculos de valores e informações complexas e não simples. Isto realmente poderia ser estatisticamente forte, e poderiam sugerir irredutibilidade. Entretanto, a simetria de um código poderia ser complexa o suficiente para não ser decodificável por nós caso a civilização seja avançada demais. Uma amostragem de razão 3 de base 37 é simples, matematicamente conhecida por nós, flexível e manipulável o suficiente para ser aplicada em diversos fenômenos que tenham o numero 3 como base e indicariam uma simples interpretação matemática de nossa própria natureza.
Conclusão
Ao que diz respeito à existência de vida inteligente no espaço e como elas possam estar ligadas ao código genético da espécie humana, ainda carece de metodologias e evidências confiáveis, dado o sinal recebido na década de 70, ou pela manifestação dos matemáticos do SETI. É fundamental estabelecer determinados conceitos e distinções em abordagens que envolvam inteligência, pois tais afirmações podem conduzir a conclusões precipitadas, especialmente de cunho religioso.
Uma contradição lógica aparente parece sempre se manter; simplicidade matemático/simétrica seria uma linguagem viável para estabelecer contato com outras civilizações inteligentes e distantes, mas poderia ser poluída por fenômenos naturais que também se encaixem nesses cálculos. Complexidade simétrica poderia indicar existência de inteligencia extraterrestre, mas talvez jamais seja interpretada por nós caso a civilização seja mais avançada.
Algo parecido ocorre com a sequência Fibonacci. Uma galáxia espiral apresenta a sequência de Fibonacci e justificaria crenças teleológicas. Esse modus operandi de atribuir valores divinos se desfaz quando se olha comparativamente fenômenos naturais que levam a formação e a quebra da sequencia Fibonacci. Por exemplo, o fato de existir galáxias irregulares, que quebra essa simetria universal de galáxias com tal sequência. O padrão espiralado das galáxias–espiral apresenta a sequencia de Fibonacci porque seu formado é garantido pela presença de um buraco negro em seu centro. Galáxias espirais têm forma de disco, com braços curvos. Aquelas com formas irregulares, ou não usuais, são conhecidas como galáxias irregulares e se originam tipicamente da disrupção pela atração gravitacional de galáxias vizinhas (Jarrett, 2007). Essas interações entre galáxias, podem ao final resultar na descaracterização do formato espiralado da galáxia, sendo assim, a sequência Fibonacci deixa de existir em tais codificações divinas também. Isso ocorre porque a sequência Fibonacci surge como um ajuste mecânico, físico natural de um fenômeno.
Alguns proponentes do DI afirmam a existência de aparente auto-organização e padrões recorrentes na natureza (como a sequência de Fibonacci dos números, espirais duplas, fractais, etc) como “prova” de design. No entanto, esses padrões podem ser mostrados para resultar de física simples (a minimização de energia potencial, quebra espontânea de simetria, autômatos celulares, etc). O algoritmo de geração de um padrão fractal é realmente muito simples e por isso não é surpresa vermos padrões fractais complexos aparecerem na natureza. Na verdade, é o que esperaríamos ver em um sistema “bottom up”, o processo de design evolucionário e da própria evolução é um tipo semelhante de método recursivo de aprimoramento de complexidade, embora não seja objetivo evolutivo. Se a descoberta de que grande parte da natureza é baseada em matemática deve ser vista como evidência contra uma entidade metafísica sobrenatural, e é um salto estranho na lógica de vincular os dois, onde a correlação não esta estabelecida.
Matemática é uma ciência exata, e uma ferramenta fundamental para a mensuração em métodos científicos. Até o momento, o que o artigo nos proporcionou, foi decifrar a matemática de abstração humana, por de trás de tal proposta. A biologia é uma ciência natural, e no caso do código genético trata de fenômenos bioquímicos, onde a vida segue processos neo-darwinianos, passeis de transformação ao longo do tempo. O estabelecimento das primeiras moléculas responsáveis pela hereditariedade e do código genético é explicado pelas ciências naturais e, única e exclusivamente por si só não necessitam de elementos exógenos para se respaldar. Independente do conceito de vida utilizado pela biologia para caracterizar a vida, ela segue um regramento geral estabelecido pelo geneticista Theodore Dobzhansky “nada em biologia faz sentido a não ser a luz da evolução”. Sendo assim, o presente trabalho não decifrou um sinal de vida inteligente fora de nosso Sistema Solar, mas sim a metodologia matemática usada para estabelecer supostos sinais de vida inteligente, e ela ainda se mostra insustentável.
Veja aqui outras pessoas que se manifestaram contra o “Wow! signal”: Intelligent signal embedded in our genetic code?, Discoveroids: Intelligent Design’s “Wow” Signal?, The “Wow! signal” of the terrestrial genetic code e The Wow! Signal: Intercepted Alien Transmission?
Referências Bibliográficas
Chris K. Caldwell, The Prime Glossary, Fermat number.
Ckrick F.H.C. The Origin of Genetic Code. Medical Research Council Laboratory of Molecular Biology Hills Road, Cambridge, England, 21 August 1968.
Cooper G, Reed C, Nguyen D, Carter M, Wang Y. Detection and formation scenario of citric acid, pyruvic acid, and other possible metabolism precursors in carbonaceous meteorites. Proc Natl Acad Sci U S A. 2011 Aug 23;108(34):14015-20.
Correa, A. L.; Silva, P. R. ; Meglhioratti , F. A; Caldeira, A. M. A. Aspectos históricos e filosóficos do conceito de vida: contribuições para o ensino de biologia. Filosofia e História da Biologia , v. 3, p. 21-40, 2008.
Darwin, C. A Origem das Espécies. Hemus – Livraria Editora Ltda, São Paulo, SP.
Dawkins R. O Gene Egoísta. Editora Itatiaia. Belo Horizonte, 2001.
E.A Kuzicheva, M.B Simakov. Abiogenic synthesis of nucleotides in conditions of space flight of the biosputnik “BION-11”. Advances in Space Research. Volume 23, Issue 2, 1999, Pages 387–391.
E.A. Kuzicheva, N.B. Gontareva. Prebiotic synthesis of nucleotides at the earth orbit in presence of lunar soil. Advances in Space Research Volume 30, Issue 6, 2002, Pages 1525–1531.
El-Hani, C. N., Videira, A. A. P. O que é vida? Para entender a biologia do século XXI. Relume Dumara. Rio de Janeiro. 2000.
Estrada M. F, Sá C. C, Queiró, J. F, Silva M, C e Costa M, J. História da Matemática. Editora: Universidade Aberta. Agosto de 2000.
Greissand S. Chin. J. Expanding the Genetic Code of an Animal. J. Am. Chem. Soc. 2011, 133, 14196–14199.
Jarrett, T. H. Near-Infrared Galaxy Morphology Atlas. California Institute of Technology. Página visitada em 09/01/2007.
King M. W. Translation of Proteins. The Medical Biochemistry Page. November 1, 2013.
Kwok, S.: Organic matter in space: from star dust to the Solar System, Astrophys. Space Sci., 319, 5–21, 2009.
Lazcano A. What Is Life? A Brief Historical Overview. Chemistry & Biodiversity – Vol. 5 (2008).
Lazcano A. Delaye L. S slas I. Becerra A. The Very Early Stages of Biological Evolution and the Nature of the Last Common Ancestor of the Three Major Cell Domains. Annu. Rev. Ecol. Evol. Syst. 2007.38:361-379.
Mendonça, A. L O.A & Videira, A. A. P. Revolução de Kuhn, Ciência hoje, vol 32, n 189 Dezembro de 2002.
Michaelian K. Thermodynamic dissipation theory for the origin of life. Earth Syst. Dynam., 2, 37–51, 2011.
Miguel Sautié Castellanos, Carlos M. Martínez Ortiz, Osmel Companioni Nápoles, José L Hernández Cáceres. El código genético desde la perspectiva de la bioinformática. Centro de Cibernética Aplicada a la Medicina, Instituto Superior de Ciencias Médicas de la Habana.
Nick Goldman, Paul Bertone, Siyuan Chen, Christophe Dessimoz, Emily M. LeProust, Botond Sipos & Ewan Birney. Towards practical, high-capacity, low-maintenance information storage in synthesized DNA. Nature (2013) doi:10.1038/nature11875.
Paulo Ribenboim. The new Book of Prime Number Records.
Primack, R. B.; Rodrigues, E. Biologia da Conservação. Editora Planta. 2001.
Ray, T. S. An Evolutionary Approach to Synthetic Biology: Zen and the Art of Creating Life. Massachusetts Institute of Technology. April 30, 2010. (doi:10.1162/artl.1993.1.1_2.179).
Ridley M. Evolution. Oxford. Third Edition. 2000.
Robson, T. Creationism and Pseudomathematics. The National Center for Science Education. Volume: 20. Issue: 4. Year: 2000.
Sagan C. Variedades da Experiência Científica. São Paulo: Companhia das Letras, 2008.
Shuch, H. P. Project ARGUS and the challenge of real-time all-sky. SETI Conference Paper, Astronomical and Biochemical Origins and the Search for Life in the Universe, IAU Colloquium 161, Publisher: Bologna, Italy, p. 693.
União Internacional de Química Pura e Aplicada. “amines”. Compêndio de Terminologia Química.
Vladimir I. Cherbaka, Maxim A. Makukov. The “Wow! signal” of the terrestrial genetic code. Icarus Volume 224, Issue 1, May 2013, Pages 228–242.
Woese, C. R.; Fox, G. E. (1977). “The concept of cellular evolution”. Journal of molecular evolution 10 (1): 1–6.
Wong JT. Coevolution theory of the genetic code at age thirty. Bioessays. 2005 Apr;27(4):416-25.
Yang, D. Peng S, Hartman, M. R, Campolongo, T. G, Rice E. J, Chang A. K, Gu, Z, Lu, G. Q, Luo. D; Enhanced transcription and translation in clay hydrogel and implications for early life evolution. Scientific Reports 3, Article number: 3165. 07 November 2013.



