
Existe uma crença amplamente difundida entre o público não-especializado de que os homens são naturalmente promíscuos, enquanto as mulheres são tímidas e exigentes. Até mesmo alguns pesquisadores – incluindo biólogos, psicólogos e antropólogos – apregoam essa noção quando interrogados sobre quase qualquer aspecto das diferenças entre machos e fêmeas, inclusive na nossa espécie. Graças a essa visão bem difundida em algumas áreas do conhecimento, certos comportamentos humanos, como o estupro, infidelidade conjugal e algumas formas de violência doméstica, foram retratados como traços adaptativos que evoluíram porque os homens são promíscuos, enquanto as mulheres são sexualmente relutantes.
Essas ideias também serviram como pedra angular para o estudo evolutivo da seleção sexual, diferenças sexuais e papéis sexuais entre os animais, alicerçados principalmente em uma hipótese conhecida como o Princípio de Bateman. Desde muito cedo, essa hipótese ganhou um status de “paradigma” nas áreas comportamentais. No entanto, apenas recentemente alguns cientistas começaram a questionar suas suposições subjacentes e o “paradigma” resultante, munidos principalmente de dados e metodologias modernas e projetos de replicabilidade.
Nesse texto, pretendo discutir a utilização do Princípio de Bateman como um paradigma vigente nas ciências comportamentais. Também pretendo discutir os projetos que buscaram testar a reprodutibilidade e replicabilidade desse estudo clássico, além da robustez dos seus resultados.

Machos promíscuos, fêmeas recatadas e do lar
Em um dos seus principais livros, The Descent of Man, Charles Darwin interpretou com base em suas observações, que “os machos de quase todos os animais têm paixões mais fortes do que as fêmeas. Consequentemente, são os machos que lutam juntos e diligentemente exibem seus encantos diante da fêmea” [1, p. 272]. Com isso, Darwin foi o primeiro a sugerir que as diferenças de tamanho e no custo presumido da energia de produção de espermatozoides versus óvulos, o que nós, biólogos, atualmente chamamos de anisogamia, seria uma possível explicação para as diferenças entre homens e mulheres no comportamento sexual [1].
Esse seu breve insight foi suficiente para que outros pesquisadores se interessassem e finalmente a expandisse para a ideia de que, como os machos produzem milhões de espermatozoides baratos, eles podem acasalar com muitas fêmeas diferentes sem incorrer em custos biológicos. Por outro lado, as fêmeas por produzirem relativamente poucos ovos, com um maior “custo energético”, deveriam ser altamente seletivas e acasalar apenas com um “melhor macho”. Além disso, esse macho escolhido forneceria esperma mais do que suficiente para fertilizar todos os óvulos femininos.

A dita primeira “explicação convincente” para as diferenças observadas na competitividade e escolha entre os sexos foi fornecida por Angus John Bateman (1919-1996) em um estudo experimental com moscas-da-fruta (Drosophila melanogaster) [2]. Um fato que merece menção é que Bateman foi um geneticista inglês que recebeu seu B.Sc. em 1940, Ph.D. em 1946 e D.Sc. em 1956 em botânica pelo King’s College, Londres, sendo afiliado a vários laboratórios durante sua carreira; trabalhando em 1948 no John Innes Horticultural Institute em Merton Park, Londres. Bateman era conhecido por pesquisar um procedimento letal dominante para testes in vivo de mutagenicidade, estudos de polinização cruzada em safras de sementes e a genética de características quantitativas em cevada e centeio, além de seus vários estudos com animais [3]. Ele publicou artigos cobrindo uma variedade de sistemas experimentais, sendo que seu trabalho de tese não envolvia estudos sobre Drosophila melanogaster, a espécie que ele usou como objeto experimental para seu agora icônico artigo de 1948. Mas ele trabalhou sob a supervisão de Ken Mathers (1911-1990), chefe do Departamento de Genética, que havia sido treinado em genética de Drosophila com o grande Thomas Hunt Morgan. A equipe de Mathers investigou vários aspectos da herança de sistemas poligênicos, usando diferentes organismos modelo (Drosophila, Petunia, Hyacinthus) [4].

Além disso, Bateman também foi membro do Partido Comunista da Grã-Bretanha durante o caso Lysenko, apresentando-se como um antilysenkoista nas convenções e dentro do Partido, enquanto escrevia em defesa de Lysenko para audiências não-partidárias [5] (claro que essa informação não é relevante para o contexto geral do texto, mas é interessante para alguns indivíduos que negam a importância de alguns autores, como o geneticista Richard Lewontin, simplesmente por sua posição política, enquanto ignora essa característica para outros).
O experimento de Bateman e sua replicabilidade
Em seu clássico experimento, Bateman aparentemente havia demostrado que o número de descendentes de uma mosca-da-fruta macho aumentava baseado no número de parceiras sexuais, enquanto uma mosca-da-fruta fêmea não ganhava um aumento no número de descendentes por acasalar com vários machos. Através desses resultados, Bateman concluiu que, como óvulos únicos são mais caros de produzir do que espermatozoides únicos, o número de descendentes produzidos por uma mosca-da-fruta fêmea era limitado principalmente por sua capacidade de produzir óvulos, enquanto o sucesso reprodutivo de um macho era limitado pelo número de fêmeas que ele inseminou. Ou seja, para Bateman o sucesso reprodutivo masculino – medido pela descendência produzida – aumentava linearmente com seu número de parceiros, mas o sucesso reprodutivo feminino atingia seu pico depois que a fêmea acasalava com apenas um macho. Ele também extrapolou suas observações e afirmou que, em nossa própria espécie, a diferença no tamanho do gameta entre os sexos resultaria em maior competição entre machos do que entre fêmeas, além de afirmar que essa característica era quase universal em todas as espécies que se reproduzem sexualmente [2].
O desenho experimental de Bateman com D. melanogaster para esse trabalho foi extremamente complexo, o que poderia ocasionar alguns vieses se não fosse bem executado. Ele montou seis séries de experimentos, com 64 populações separadas, cada uma em um pequeno frasco, permitindo-lhe monitorar o número de descendentes para um total de 440 moscas individuais (220 de cada sexo, sendo que [6] é dito que eram 215 indivíduos de cada sexo). Além disso, essas 64 populações foram organizadas em seis “séries” que variaram no número de repetições, no número de dias em que o acasalamento poderia ocorrer, no número de machos e fêmeas nas populações, nas idades de machos e fêmeas e em suas mutações marcadoras dominantes. Bateman tratou cada série como um experimento separado, embora não tenha sido explícito sobre seus motivos, sendo essa uma informação metodológica importante para se entender o experimento [2].
Cada mosca de sua primeira geração (F1) era um heterozigoto dominante, isto é, o portador de uma mutação fenotípica individual em um alelo junto com um alelo de tipo selvagem no locus marcador. A identificação de Bateman de quais moscas experimentais produziram descendentes foi baseada não em observações comportamentais, mas em inferências indiretas de mutações visíveis de um único gene transmitidas por descendentes de mosca. Ele nunca realmente monitorou o comportamento das moscas, com que frequência as moscas se acasalaram (ou não) ou com quantos parceiros cada uma delas copulou. Com isso, aqui já podemos pontuar a primeira crítica metodológica ao experimento de Bateman. Vale lembrar que embora Bateman não tenha registrado o comportamento de machos e fêmeas em suas populações experimentais, ele concluiu a priori, como Darwin havia hipotetizado (1871), que a competição macho-macho e a escolha feminina eram as causas da alta variação na aptidão masculina.
Tang-Martínez e Ryder [7], ao analisar a metodologia de Bateman, afirmaram que, sem dúvida, esse desenho experimental era “engenhoso e elegante”, especialmente “em uma época em que as técnicas moleculares não estavam disponíveis”, porém “era indireto e potencialmente falho”. Da mesma forma, Gowaty e colaboradores [8], ao tentar replicar os resultados de Bateman, demonstraram a “falha fatal” no desenho experimental de Bateman, que poderia fornecer uma amostra distorcida ao estudo – sendo assim, um enorme viés para qualquer conclusão obtida posteriormente. No estudo intitulado “Nenhuma evidência de seleção sexual em uma repetição do estudo clássico de Drosophila melanogaster de Bateman”, Gowaty e colaboradores [8] demonstraram que “o método de Bateman superestimou sujeitos com zero parceiros, subestimou sujeitos com um ou mais parceiros e produziu estimativas sistematicamente tendenciosas do número de descendentes por sexo”. Esse mesmo estudo concluiu que: “a metodologia de Bateman avaliou erroneamente as variações de aptidão que são as variáveis-chave da seleção sexual”.
Desde o desenho experimental, Bateman assumiu a priori que as frequências dos fenótipos parentais na prole seguiriam as expectativas de Mendel: um quarto seria duplo-mutante que herdaria o gene dominante de cada progenitor, sendo essa a prole a partir da qual Bateman contou o número de parceiros por criador (M♀ M♂); metade da descendência seria mutante único, herdando o gene dominante de um dos pais e o alelo de tipo selvagem do outro pai (M♀/♂ W♀/♂); e um quarto não herdaria nenhuma das mutações marcadoras de seus pais (W♀ W♂). Essa seria a expectativa padrão em um quadro de Punnett de seus cruzamentos.
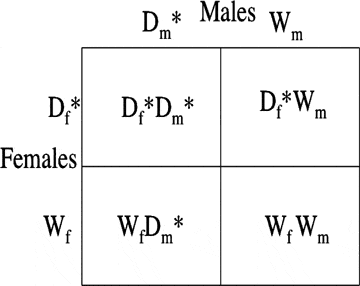
Com base nesses pressupostos, também podemos confirmar que a suposição crucial do método de Bateman é que não haveria redução da viabilidade da prole a partir da herança de marcadores parentais, particularmente quando a prole herdava uma mutação de cada um de seus pais (M♀ M♂), sendo que ao não registrar o comportamento de machos e fêmeas em suas populações experimentais, essa prole seria a única da qual a variância do número de parceiros para cada adulto poderia ser inferida usando o método proposto por Bateman [8]. No entanto, voltamos para mais um problema crucial no experimento, “a viabilidade completa de todos os genes marcadores” assumida por Bateman [2, p. 353] era um critério que estava longe de ser demonstrado e o próprio autor observou que a maioria de seus genes marcadores eram letais na condição de homozigoto. Com isso, podemos afirmar que Bateman tinha conhecimento dessas falhas potenciais e que passou a estimar o sucesso reprodutivo de cada sujeito mesmo com esse enorme viés metodológico.
No entanto, ao replicar seus experimentos, Gowaty e colaboradores [8] descobriram que as moscas com duas mutações (M♀ M♂) teriam uma menor probabilidade de sobreviver até a idade adulta. Um dos principais motivos é que essas moscas utilizam suas asas não apenas para pairar, mas também durante a cópula, razão pela qual asas encaracoladas apresentam uma grande desvantagem (e seria esperado que esse fator tivesse sido apontado ao longo dos anos em que o experimento foi citado na comunidade científica). Os espécimes com olhos deformados podem ter uma sobrevivência ainda mais difícil. Os 25% da prole nascida com ambas as mutações tinham ainda uma probabilidade maior de morrer antes de serem contadas por Bateman ou Gowaty. Nesse estudo, Gowaty também observou que a fração de descendentes com mutantes-duplos (M♀ M♂) estava significativamente abaixo dos 25% esperados, o que significava que Bateman não teria sido capaz de quantificar com precisão o número de parceiros para cada sujeito adulto. Além disso, os autores observaram que a metodologia de Bateman resultou em mais descendentes atribuídos aos pais do que às mães, algo que é impossível quando cada descendente deve ter um pai e uma mãe. Por último, mas ainda extremamente relacionado, Gowaty verificou que as proporções de descendência em cada uma das séries encontradas pela replicação “afastou-se fortemente das expectativas de Mendel”.
Bateman concluiu em seu artigo que [2 p. 360]: “enquanto apenas 4% das fêmeas não estavam representadas na progênie, 21% dos homens não estavam representados”. A proporção de 4:21 foi interpretada como mostrando que os machos tinham cerca de cinco vezes mais probabilidade do que as fêmeas de perder na competição reprodutiva. Bateman começou seu estudo com a convicção de que “a variação na fertilidade é, na verdade, uma medida da intensidade da seleção” [2 p. 353]. Consequentemente, ele acreditava que seus resultados demonstravam que a variação no sucesso da produção de descendentes fornecia evidências para diferentes níveis de intensidade de seleção. Em resumo, Bateman concluiu que os machos das moscas-da-fruta produziam muito mais descendentes viáveis quando têm vários parceiros, mas que as fêmeas produziam o mesmo número de filhos adultos, quer tenham um ou vários parceiros. No entanto, graças a diversos problemas metodológicos, Gowaty e colaboradores demonstraram que os dados apresentados por Bateman eram decididamente inconclusivos.
Em seu projeto de replicabilidade, Gowaty e colaboradores demonstraram de forma empírica que os dados de Bateman falharam em corresponder a uma suposição fundamental de atribuições genéticas de parentesco, sendo que os marcadores usados para identificar os sujeitos individuais estavam influenciando os parâmetros medidos, como o número de parceiros e o número de filhos. Quando a prole morre de mutações herdadas do marcador, os resultados se tornam tendenciosos, indicando que o método é incapaz de abordar de forma confiável a relação entre o número de casais e o número de descendentes. Com isso, como citamos anteriormente, o método de Bateman superestimou indivíduos com zero parceiros, subestimou indivíduos com um ou mais parceiros e produziu estimativas sistematicamente tendenciosas do número de descendentes por sexo, avaliando assim erroneamente as variações de aptidão que são as variáveis-chave da seleção sexual [4,6,8].
As seis séries amostrais
Como citei anteriormente, as 64 populações analisadas por Bateman foram organizadas em seis “séries” amostrais que variaram no número de repetições, no número de dias em que o acasalamento poderia ocorrer, no número de machos e fêmeas nas populações, nas idades de machos e fêmeas e em suas mutações marcadoras dominantes. Além disso, Bateman tratou cada série como um experimento separado, embora não tenha sido explícito sobre seus motivos [2].
Cada uma das 64 populações de Bateman consistia em cinco fêmeas e cinco machos ou três fêmeas e três machos. Bateman permitiu que eles se acasalassem e ovipositassem por alguns dias, mas o número de dias variou. Durante o experimento, Bateman controlou o efeito dos diferentes marcadores genéticos, o efeito da idade e ancestralidade genética, no entanto, algumas séries implicaram em cruzamentos recíprocos (inversão dos sexos) [4].
Ao analisar seus resultados, Bateman afirmou que “a diferença de sexo na variância da fertilidade não se deve à maior sensibilidade dos machos a mudanças classificáveis, ambientais ou genéticas. A pista para essa diferença de sexo deve ser buscada no próprio mecanismo da reprodução sexual” [2 p. 359]. No entanto, enquanto ele estava analisando e reunindo seus dados, notou “que as séries 5 e 6 diferiam um pouco do resto” [2 p. 361].
Com essa observação, Hoquet [4] destaca que Bateman resolveu combinar as séries 1 e 4, assim como as séries 5 e 6. Da mesma forma, Hoquet [4] destaca que a decisão de encapsular seus resultados em dois gráficos diferentes apresentados como Figura 1 [2, p. 362] também foi posterior à obtenção dos primeiros resultados e que ambos os gráficos (a) e (b) apresentam uma relação linear entre o número de companheiros e o número de descendentes para os machos. Além disso, destaca que um efeito semelhante para o sexo feminino, embora em menor grau, também é evidente, especialmente no gráfico da Fig. 1a. e que apenas no gráfico da Fig. 1b parece haver um platô após um parceiro para as fêmeas.
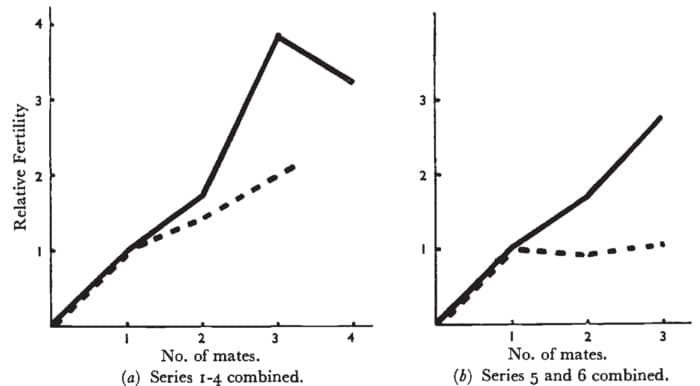
É bom destacar que esse gráfico será extremamente importante na segunda parte desse texto.
Conclusões parciais
Terminamos por aqui a primeira parte deste texto.
Um fato que merece ser destacado é que esse texto não tem como objetivo principal ser uma critica puramente à Bateman e que precisamos lembrar que ele realizou seu trabalho sem ferramentas computacionais modernas ou mesmo metodologias biomoleculares atuais, utilizando uma abordagem inovadora para sua época. Ele tentou testar empiricamente a hipótese de Darwin e buscou enfatizar a importância da variação de aptidão para a seleção sexual. No entanto, não podemos ignorar as falhas metodológicas em um experimento considerado clássico e modelo em uma determinada área do conhecimento, principalmente em um período em que a Crise de Reprodutibilidade é um tema em voga. Além disso, os estudos de replicabilidade de Bateman levantam um questionamento extremamente importante sobre o que consideramos como uma evidência positiva para a seleção sexual.
O questionamento de Gowaty e colaboradores na conclusão do seu artigo também deve ser destacado e uma reflexão realizada: “como os leitores anteriores não conseguiram identificar os problemas inferenciais com o estudo original de Bateman?”. Afinal, o trabalho de Bateman é um artigo clássico que foi lido por legiões de estudantes de pós-graduação (além de pesquisadores seniores), sendo esses competentes o suficiente para verificar esses erros. Sem contar que os resultados de Bateman são apresentados em vários livros didáticos de biologia, e seu artigo vem sendo citado até hoje em um número expressivo de outros estudos científicos. No entanto, grande parte desses novos estudos ignoram todas as críticas metodológicas apresentadas por Gowaty e colaboradores, Hoquet, entre outros.
Além disso, os resultados de Bateman foram aceitos com tanto entusiasmo no meio científico que o artigo caracterizou ao longo do tempo o que vale e o que não vale a pena investigar na biologia do comportamento. Como veremos na segunda parte do texto, seus resultados geram os chamados Princípios de Bateman que nivelaram diversas pesquisas de seleção sexual. Além disso, mesmo sem ter sido validado, sua aceitação o estabeleceu como um paradigma dentro da área comportamental [3].
A replicabilidade de estudos científicos é um princípio da ciência, razão pela qual a metodologia de Bateman deveria ter sido replicada assim que se tornou importante na década de 1970, com a redescoberta de seu trabalho por Trivers [9]. É nesse momento que falaremos sobre fraudes, ética em pesquisa e os problemas envolvendo a ascensão de Bateman como um paradigma.
Para concluir essa parte do texto, deixo uma última parte da conclusão de Gowaty e colaboradores:
O poder paradigmático da visão de mundo capturado nas conclusões de Bateman e a frase “Princípios de Bateman” pode deslumbrar os leitores, obscurecendo da vista as fraquezas metodológicas e hipóteses alternativas razoáveis…
Observação
Essa é a primeira parte de um texto previsto para ter duas partes. Na próxima, discutiremos o redescobrimento de Bateman, seus princípios e seu estabelecimento como paradigma.
Referências
- Darwin, C. (1871). The descent of man, and selection in relation to sex. John Murray. London, England. UK, 1, 423.
- Bateman, A. J. (1948). Intra-sexual selection in Drosophila. Heredity, 2(3), 349-368.
- Debus, A. G. (1968). World who’s who in science (p. 490). Chicago: Marquis-Who’s Who.
- Hoquet, T. (2020). Bateman (1948): rise and fall of a paradigm?. Animal Behaviour.
- Paul, D. B. (1983). A war on two fronts: JBS Haldane and the response to Lysenkoism in Britain. Journal of the History of Biology, 16(1), 1-37.
- Snyder, B. F., & Gowaty, P. A. (2007). A reappraisal of Bateman’s classic study of intrasexual selection. Evolution: International Journal of Organic Evolution, 61(11), 2457-2468.
- Tang-Martinez, Z., & Ryder, T. B. (2005). The problem with paradigms: Bateman’s worldview as a case study. Integrative and Comparative Biology, 45(5), 821-830.
- Gowaty, P. A., Kim, Y. K., & Anderson, W. W. (2012). No evidence of sexual selection in a repetition of Bateman’s classic study of Drosophila melanogaster. Proceedings of the National Academy of Sciences, 109(29), 11740-11745.
- Trivers, R. L. (1972). Parental investment and sexual selection. In ‘Sexual Selection and the Descent of Man’.(Ed. B. Campbell.) pp. 136–179. Aldinc: Chicago, 13(2).



